
約1年半前、私はHabrで、脳がどのように機能するか、人工知能を作成するための可能な方法は何かというビジョンとともに、 ビデオ講義のサイクルを作成しました。 それ以来、大きな進歩を遂げることができました。 何かがより深く理解されることが判明しました。何かはコンピューター上でシミュレートすることができました。 素晴らしいことは、プロジェクトの作業に積極的に関与している志を同じくする人々がいることです。
この一連の記事では、現在取り組んでいる知性の概念について話し、脳の仕事のモデリングの分野で根本的に新しいいくつかのソリューションを実証する予定です。 しかし、ストーリーを理解し、一貫性を保つために、新しいアイデアの説明だけでなく、脳全体の機能に関するストーリーも含まれます。 特に最初の段階では、いくつかのことは単純で有名に思えるかもしれませんが、ストーリーの一般的な証拠を大きく決定するため、それらをスキップしないことをお勧めします。
脳の一般的な考え方
神経細胞、それらはニューロンであり、信号を伝達する繊維と共に神経系を形成します。 脊椎動物では、大部分のニューロンは頭蓋腔と脊柱管に集中しています。 これは中枢神経系と呼ばれます。 したがって、脳と脊髄はその成分として区別されます。
脊髄は体内のほとんどの受容体から信号を収集し、脳に伝達します。 視床の構造を通して、それらは大脳半球の皮質に分布して投影されます。

皮質への情報の投影
大脳半球に加えて、実際には小さな独立した脳である小脳も情報の処理に関与しています。 小脳は、すべての動きの正確な運動性と協調性を提供します。
視覚、聴覚、嗅覚は脳に外界に関する情報の流れを提供します。 パスに沿って通過するこのストリームの各コンポーネントも皮質に投影されます。 皮質は、脳の外表面を構成する厚さ1.3〜4.5 mmの灰白質の層です。 折り畳みによって形成される畳み込みにより、樹皮は、平らな形よりも面積が3倍小さくなるように梱包されます。 1つの半球の総地殻面積は約7000平方Cmです。
その結果、すべての信号が皮質に投影されます。 投射は、皮質の限られた領域に分布している神経線維の束によって行われます。 外部情報が投影される領域、または脳の他の部分からの情報は、皮質のゾーンを形成します。 そのようなゾーンに送られる信号に応じて、独自の専門性があります。 皮質の運動ゾーン、感覚ゾーン、ブロカ、ウェルニッケ、視覚ゾーン、後頭葉を区別するのは、約100種類のゾーンだけです。
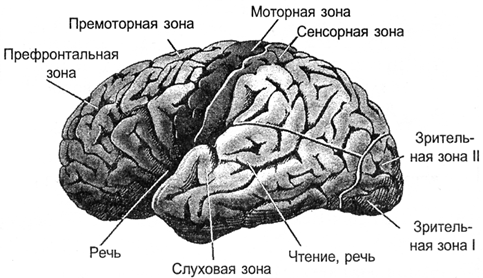
樹皮のゾーン
垂直方向では、皮質は通常6つの層に分割されます。 これらのレイヤーには明確な境界はなく、1つまたは別のタイプのセルの優位性によって決定されます。 皮質のさまざまなゾーンでは、これらのレイヤーをさまざまな方法で表現できます。 しかし、一般的に、皮質は非常に普遍的であると言え、その異なるゾーンの機能は同じ原理に従うと仮定しています。

樹皮の層
求心性線維を介して、信号が皮質に入ります。 それらは皮質のIII、IVレベルに到達し、そこで求心性線維が落ちた場所の近くにあるニューロンに分布します。 ほとんどのニューロンは、皮質内に軸索結合を持っています。 しかし、一部のニューロンにはそれを超える軸索があります。 これらの遠心性繊維を介して、信号は脳の外側、たとえば実行器官に送られるか、または他の半球の皮質の他の部分に投射されます。 信号伝送の方向に応じて、遠心性ファイバーは通常次のように分類されます。
- 1つの半球の皮質の別々のセクションを接続する結合繊維。
- 2つの半球の皮質を接続する交連線維。
- 皮質を中枢神経系の下部の核に接続する投射線維。
皮質の表面に垂直な方向をとると、この方向に沿って位置するニューロンが同様の刺激に反応することがわかります。 このような垂直に配置されたニューロンのグループは、一般に皮質列と呼ばれます。
大脳皮質は、別々のゾーンにカットされた大きなキャンバスとして想像できます。 各ゾーンのニューロンの活動の画像は、特定の情報をエンコードします。 皮質ゾーンを超えて伸びる軸索によって形成された神経線維の束は、投射接続のシステムを形成します。 特定の情報が各ゾーンに投影されます。 さらに、自身のゾーンと反対側の半球の両方から来ることができるいくつかの情報フローは、1つのゾーンに同時に到着できます。 情報の各ストリームは、神経束の軸索の活動によって描かれた一種の絵のようなものです。 皮質の別のゾーンの機能は、多くの投影の取得、情報の保存、処理、独自のアクティビティの画像の形成、さらにこのゾーンの作業から得られる情報の投影です。
脳のかなりの部分が白質です。 それは、同じ投影経路を作成するニューロンの軸索によって形成されます。 下の図では、白質は、皮質と脳の内部構造の間の光の充満として見ることができます。
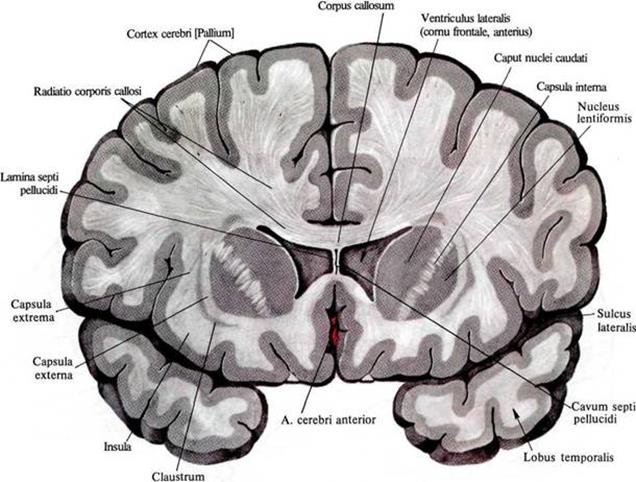
脳の前頭部における白質の分布
拡散スペクトルMRIを使用して、個々の繊維の方向を追跡し、皮質ゾーンの接続性の3次元モデルを構築することができました(Connectomicsプロジェクト(Connect))。
下の写真は、債券の構造の良い考えを示しています(Van J. Wedeen、Douglas L. Rosene、Ruopeng Wang、Guangping Dai、Farzad Mortazavi、Patric Hagmann、Jon H.Kaas、Wen-Yih I. Tseng、2012年)。

左半球からの眺め

背面図

右の眺め
ちなみに、左右の半球の投影経路の非対称性は、背面図ではっきりと見えます。 この非対称性は、学習時に半球が獲得する機能の違いを大きく決定します。
ニューロン
脳の基底はニューロンです。 当然、ニューラルネットワークを使用した脳のモデリングは、その動作の原理は何かという質問に対する答えから始まります。
実際のニューロンの仕事は化学プロセスに基づいています。 ニューロンの内部環境と外部環境の間には、電位差があります-膜電位は約75ミリボルトです。 それはナトリウム-カリウムポンプのように働く特別なタンパク質分子の働きにより形成されます。 ATPヌクレオチドのエネルギーにより、これらのポンプはカリウムイオンを細胞の内側に、ナトリウムイオンを細胞の外側に駆動します。 この場合のタンパク質はATPアーゼ、すなわちATPを加水分解する酵素として作用するため、「ナトリウムカリウムATPアーゼ」と呼ばれます。 その結果、ニューロンは、内部が負電荷、外部が正電荷の充電済みコンデンサに変わります。

ニューロンスキーム(マリアナルイスビジャレアル)
ニューロンの表面は分岐プロセスで覆われています-樹状突起。 他のニューロンの軸索終末は樹状突起に隣接しています。 それらの接続の場所はシナプスと呼ばれます。 シナプス相互作用により、ニューロンは着信信号に応答し、特定の状況下で、スパイクと呼ばれる独自のインパルスを生成できます。
シナプスの信号伝達は、神経伝達物質と呼ばれる物質が原因で発生します。 軸索に沿った神経インパルスがシナプスに入ると、このシナプスに特有の神経伝達物質分子が特別な小胞から放出されます。 シグナルを受け取るニューロンの膜には、タンパク質分子-受容体があります。 受容体は神経伝達物質と相互作用します。
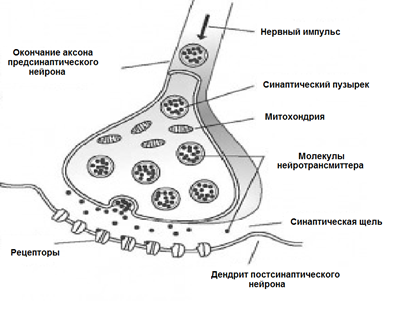
化学シナプス
シナプス間隙に位置する受容体はイオンチャネル型です。 この名前は、イオンを輸送できるイオンチャネルでもあるという事実によって強調されています。 神経伝達物質は受容体に作用するため、イオンチャネルが開きます。 したがって、膜は、どのチャネルが影響を受け、それに応じてどのタイプのシナプスに依存するかに応じて、脱分極または過分極されます。 興奮性シナプスでは、陽イオンを細胞に通過させるチャネルが開き、膜が脱分極します。 抑制性シナプスでは、陰イオンを伝導するチャネルが開き、膜の過分極につながります。
特定の状況では、シナプスはその感受性を変化させる可能性があり、これはシナプス可塑性と呼ばれます。 これは、1つのニューロンのシナプスが外部信号に対する異なる感受性を獲得するという事実につながります。
同時に、多くの信号がニューロンのシナプスに到着します。 抑制性シナプスは、ケージ内の電荷の蓄積に向かって膜電位を引き出します。 それどころか、シナプスを活性化すると、ニューロンを放電させようとします(下図)。
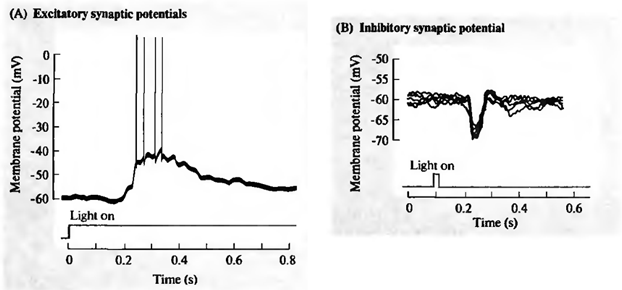
網膜神経節細胞の興奮(A)および阻害(B)(Nicholls J.、Martin R.、Wallas B.、Fuchs P.、2003)
合計アクティビティが開始しきい値を超えると、活動電位またはスパイクと呼ばれる放電が発生します。 スパイクは、電気刺激を生成するニューロン膜の急激な脱分極です。 パルス生成のプロセス全体は約1ミリ秒続きます。 同時に、パルスの持続時間も振幅も、それを引き起こした原因がどれほど強いかに依存しません(下図)。

神経節細胞の活動電位の登録(Nicholls J.、Martin R.、Wallas B.、Fuchs P.、2003)
はんだ付け後、イオンポンプは神経伝達物質の再取り込みを行い、シナプス間隙をきれいにします。 スパイクに続く不応期の間、ニューロンは新しいインパルスを生成できません。 この期間の持続時間は、ニューロンが可能な最大生成頻度を決定します。
シナプスでの活動の結果として生じる癒着は、誘発と呼ばれます。 誘発されたスパイクの繰り返し率は、入力信号がニューロンシナプスの感度設定とどれだけ一致するかをエンコードします。 着信信号がニューロンを活性化する敏感なシナプスに正確にあり、これが抑制性シナプスに来る信号と干渉しない場合、ニューロンの反応は最大になります。 このような信号によって記述される画像は、ニューロンの刺激特性と呼ばれます。
もちろん、ニューロンの働きの概念は単純化すべきではありません。 一部のニューロン間の情報は、スパイクによって送信されるだけでなく、細胞内の内容を接続し、電位を直接送信するチャネルによっても送信されます。 この伝播は段階的と呼ばれ、接続自体は電気シナプスと呼ばれます。 ニューロンの体までの距離に応じた樹状突起は、近位(近い)と遠位(遠い)に分けられます。 遠位樹状突起は、半自律的な要素として機能するセクションを形成できます。 興奮のシナプス経路に加えて、代謝型癒着を引き起こすシナプス外メカニズムがあります。 誘発された活動に加えて、自発的な活動もあります。 そして最後に、脳ニューロンはグリア細胞に囲まれており、グリア細胞も進行中のプロセスに大きな影響を及ぼします。
長い進化のパスは、脳がその仕事で使用する多くのメカニズムを作成しました。 それらのいくつかはそれ自体で理解することができ、他の意味は十分に複雑な相互作用を考慮する場合にのみ明らかになります。 したがって、ニューロンの上記の説明を網羅的なものと見なさないでください。 より深いモデルに進むには、まずニューロンの「基本的な」特性を扱う必要があります。
1952年、アラン・ロイド・ホジキンとアンドリュー・ハクスリーは、巨大なイカの軸索における神経信号の生成と伝達を決定する電気的メカニズムの説明をしました(ホジキン、1952年)。 1963年にノーベル医学生理学賞で評価されたもの。 Hodgkin-Huxleyモデルは、常微分方程式系によってニューロンの動作を記述します。 これらの方程式は、アクティブな媒体での自動波プロセスに対応します。 それらは、多くのコンポーネントを考慮に入れます。各コンポーネントは、実際の細胞内に独自の生物物理学的アナログを持っています(下図)。 イオンポンプは電流源I pに対応します。 細胞膜の内側の脂質層は、C mの容量を持つコンデンサーを形成します。 シナプス受容体のイオンチャネルは、時間tで変化する供給信号と膜電位Vの合計値に依存する導電率g nを提供します。膜孔の漏れ電流は導体g Lを作成します。 イオンチャネルを通るイオンの移動は、電気化学的勾配の影響下で発生します。電気化学的勾配は、起電力E nおよびE Lを持つ電圧源に対応します。
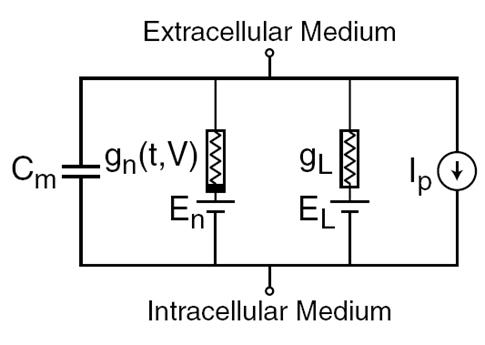
Hodgkin-Huxleyモデルの主要コンポーネント
当然、ニューラルネットワークを作成するとき、ニューロンモデルを単純化し、その中に最も重要なプロパティのみを残したいという要望があります。 最も有名で人気のある単純化されたモデルは、1940年代初期に開発されたMcCulloch – Pitts人工ニューロンです(McCulloch J.、Pitts W.、1956)。
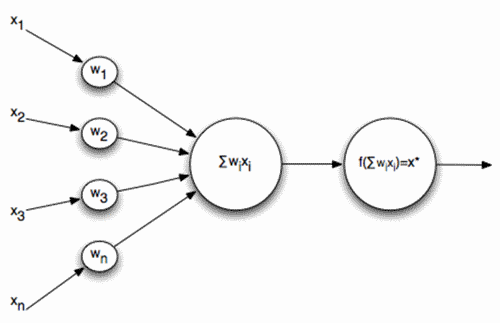
マカロック-ピッツフォーマルニューロン
そのようなニューロンの入力に信号が送信されます。 これらの信号は重み付けされて要約されます。 さらに、シグモイドなどの特定の非線形活性化関数がこの線形結合に適用されます。 多くの場合、シグモイドとしてロジスティック関数を使用します。
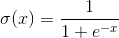
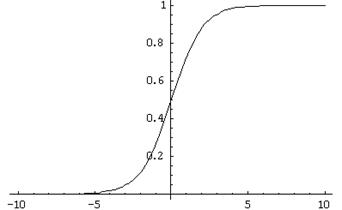
ロジスティック関数
この場合、正式なニューロンのアクティビティは次のように記述されます。
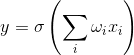
その結果、このようなニューロンはしきい値加算器になります。 かなり急なしきい値関数では、ニューロンの出力信号は0または1のいずれかです。入力信号とニューロンの重みの加重和は、2つの画像の畳み込みです:入力信号の画像とニューロンの重みで記述される画像。 畳み込みの結果が高いほど、これらの画像の対応が正確になります。 つまり、実際には、ニューロンは、配信された信号がシナプスに記録された画像にどれだけ似ているかを決定します。 畳み込み値が特定のレベルを超え、しきい値関数が単一に切り替わると、これは提示された画像を認識したというニューロンによる決定的なステートメントとして解釈できます。
実際のニューロンは、何らかの方法でマカロック-ピッツニューロンに似ています。 それらのスパイクの振幅は、それらを引き起こしたシナプスでの信号に依存しません。 スパイクがあるかどうか。 しかし、実際のニューロンは単一のインパルスではなく、インパルスシーケンスで刺激に応答します。 この場合、パルス周波数が高いほど、ニューロンの画像特性がより正確に認識されます。 つまり、このようなしきい値加算器からニューラルネットワークを構築し、静的入力信号を使用すると、何らかの出力結果が得られますが、この結果は実際のニューロンの動作を再現するにはほど遠いということです。 ニューラルネットワークを生物学的プロトタイプに近づけるには、時間パラメータを考慮して信号の周波数特性を再現し、ダイナミクスで作業をシミュレートする必要があります。
しかし、あなたは他の方法で行くことができます。 たとえば、ニューロンの活動の一般化された特性を区別できます。これは、インパルスの頻度、つまり特定の期間のスパイクの数に対応します。 この説明に進むと、ニューロンを単純な線形加算器と考えることができます。
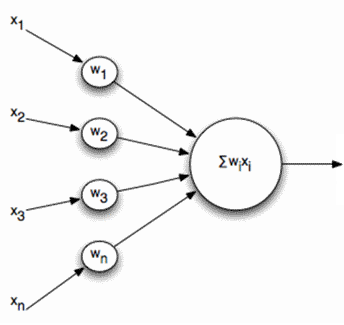
線形加算器
そのようなニューロンの出力信号とそれに応じた入力信号は、2原子(0または1)ではなく、特定のスカラー量で表されます。 アクティベーション関数は次のように記述されます
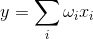
線形加算器は、パルスニューロンとは根本的に異なるものとして認識されるべきではありません。モデリングまたは記述するときに、より長い時間間隔に移動することができます。 インパルスの記述はより正確ですが、線形加算器への移行は多くの場合、モデルの強力な単純化によって正当化されます。 さらに、パルスニューロンでは見にくいいくつかの重要な特性は、線形加算器にとっては明らかです。
継続
中古文学
アレクセイ・レドズボフ (2014)