理論のビット
複雑なネットワークは、重要なトポロジ特性を持つグラフと呼ばれます。 非自明なトポロジプロパティとは、通常、グラフのノード間の接続がcな法則に従って分散され、さまざまなクラスター構造、ハブ、脆弱性などが与えられることを意味します。
複雑なネットワークの概念は、前世紀の50年代の科学作品の引用を追跡するというアイデアから生まれ、徐々に成長し、インターネット、ソーシャルネットワーク、道路ネットワーク、さらには理論物理学の多くの分野、さらには量子さえも取り込みました。 複雑なネットワークの先駆者は、ハンガリーの数学者ポール・エルデスでした。彼は、ネットワーク理論と関連する問題に関する多数の記事を発表しています。 科学界での彼の栄誉において、引用が冗談めかしてエルドシュ数で測定されたことは興味深い。 これは、Erdoshと共同研究者のネットワーク内の別の著者との間の接続の長さを示すコミックメトリックです。 つまり、Vasily PupkinがAlfred Renyiの共著者であり、その共著者がErdoshの共著者である場合、VasilyのErdosh番号は2になります(ポールとアルフレッドにはそれぞれ0と1があります)。
複雑なネットワークの理論における重要な問題(およびその結果としての特性)の1つは、ネットワークをクラスターに分割する問題です。 クラスターは、ネットワークの残りの部分よりも相互に接続されているノードの配列です。 現在、クラスタリングアルゴリズムは数百ではないにしても数十ありますが、すべてに制限と欠点があります:それらは遅すぎるか、計算時間がノードの数とともに天文学的に増加するか、初期タスクの条件に応じて精度が低下します。 この記事では、勾配クラスターアルゴリズムを使用します。その本質は次のとおりです。各ノードについて、他のノードとの最大接続のみを残し(したがって、自己接続は除外します)、他のすべてを削除します。 したがって、ネットワークはサブネットに分割され、サブネットになります。
上の図では、クラスターは目で簡単に見つけられますが、下の図ではわかりますか?


申込み
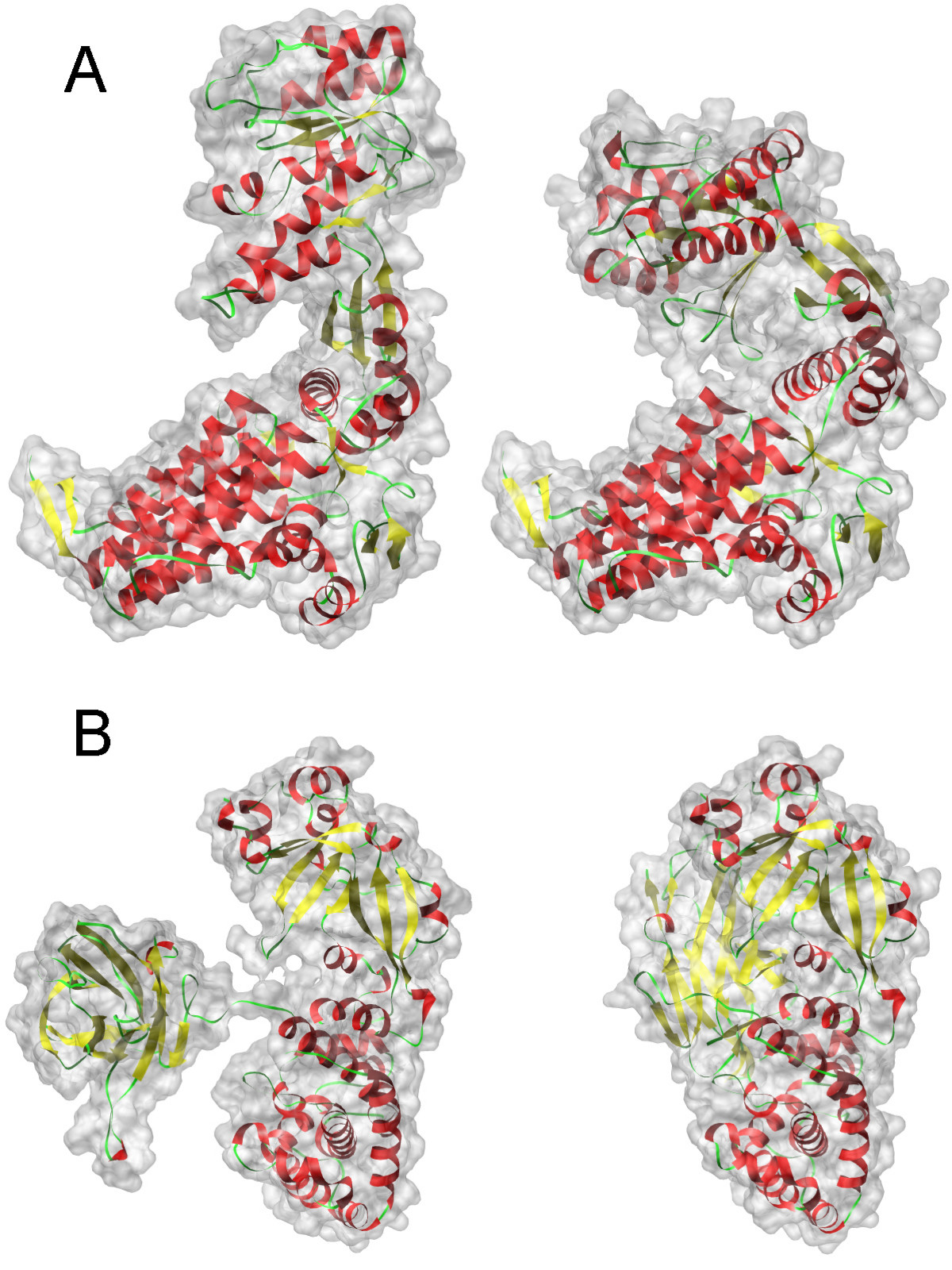
なぜこれがすべて必要なのですか? 実際には、外部要因(温度、圧力、イオン、水など)の影響下にあるタンパク質は、さまざまな状態(立体配座)にある可能性があり、さまざまな生物学的機能に対応します。 通常、状態は、慣性半径、水素結合の数、特定の原子間の距離など、特定のパラメーターまたはいくつかのパラメーター(順序パラメーター)で記述されます。
写真は、2つの異なるタンパク質の2つの状態を示しています。 それぞれ異なる状態の関数は異なります
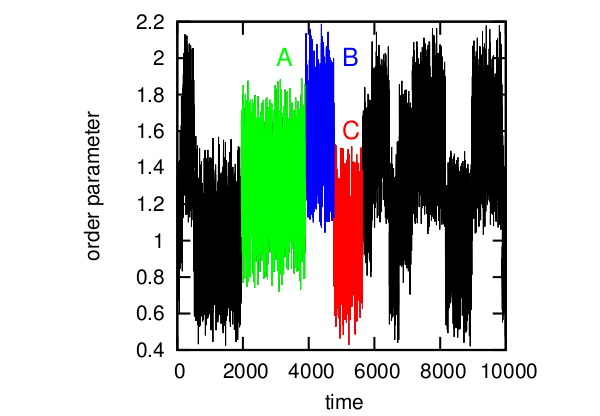
最も単純な例を考えてみましょう。 タンパク質の条件付き順序パラメーターの時系列。 ここでは、タンパク質の3つの状態に対応する3つの状態(A、B、C)を区別できます。 通常、このような場合、式F = -kT log(P)が使用されます。ここで、kTは定数、Pは状態の確率、Fは自由エネルギーであり、最小エネルギーが異なるタンパク質構造に対応する自由エネルギープロファイルが構築されます。 さらに、外部要因に応じて、システムは複数のエネルギー井戸間をジャンプすると言われているようです。 すべてがうまくいくようです。
時系列の例。
自由エネルギープロファイル。
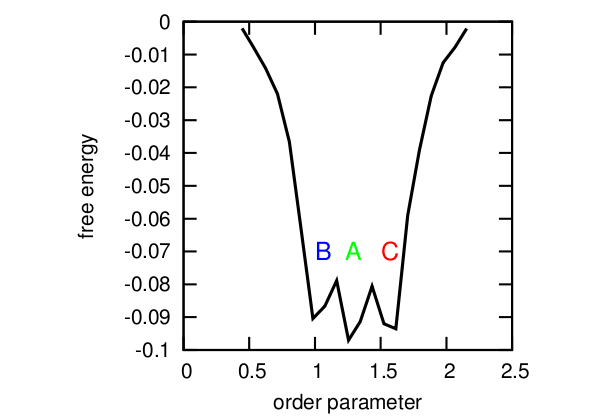
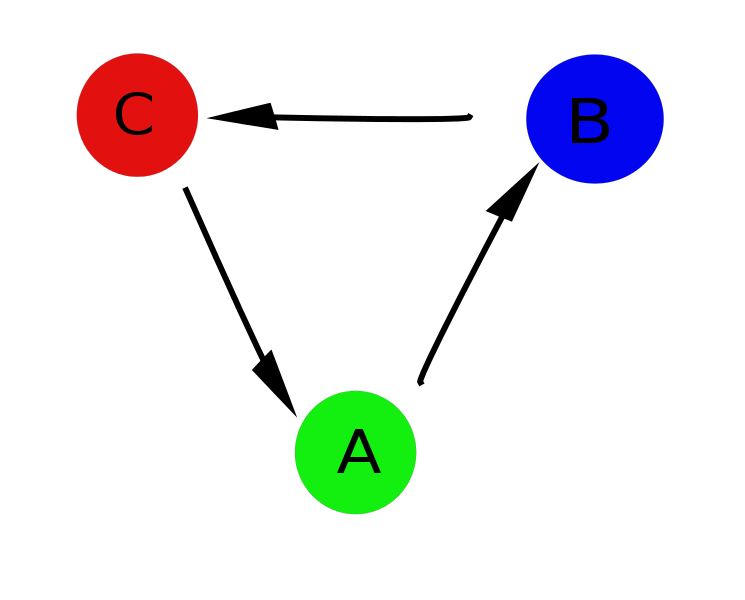
しかし、いくつかの問題があります:最初の、そして最も明白な-どのパラメーター値がどの状態に対応するかを言うことが常に可能とは限りません(たとえば、OP = 1.4では、3つの状態すべてが可能です)、そして得られたプロファイルは実際の画像をわずかにゆがめます。 第二に、実際には、厳密に定義されたサイクルA-> B-> Cがあり、状態Aから状態Cへの遷移は直接不可能であり、結果としてすべてを1つの軸に投影するため、現実。
そして、ここでネットワークが助けになります。 各瞬間の順序パラメーターの値とネットワークノードの間の対応を設定し、時間的に隣接する2つの間の条件単位に等しい重みを持つ接続を作成できます。分析中にこの瞬間が繰り返される場合は、それに応じて重みを増やします。 さらに、既に推測したように、上記のクラスタリングアルゴリズムを適用し、システムを実際に説明する図を取得します。