数か月前に、一連の記事が「Logic of Thinking」という一般的なタイトルで公開されました。 楽観的には、1〜2か月で継続する予定でした。 しかし、人生は独自の調整を行いました。 皮質のパターン波モデルをモデリングすると、Habrのサイクルの続きを書くなど、他のすべてをしばらく延期しなければならないという興味深い結果が得られました。
しかし、それほど前ではありませんが、プレプリント( http://arxiv.org/abs/1406.6901 )に記事を書いて投稿しました。 ある意味では、以前は波のモデルに興味を持っていた人にとっては興味深いかもしれません。 モデルの重要なポイントは、ニューロンがシナプスの重みで記述された1つの画像だけでなく、この画像とは異なる他の多数の信号も記憶および認識できるということです。 もちろん、ニューロンのこのような複雑さは多くの既存の理論に反しており、深刻な正当化以上のものを必要とします。 以下では、私のモデルを支持して、記事で提示された議論の1つを説明しようとします。
この記事は、サイクルの継続と見なされるべきではなく、むしろ前編です。 私自身、これを樹状突起棘の重要な役割の議論と呼びました。
よく知られているものを繰り返すことから始めましょう。
ニューロンの内部環境と外部環境の間には、電位差があります-膜電位は約70ミリボルトです。 イオンポンプのような働きをするタンパク質分子によって形成されます。 その結果、膜は分極し、負の電荷が細胞内に蓄積し、正の電荷が外部に蓄積します。
ニューロンの表面は分岐プロセスで覆われています-樹状突起。 他のニューロンの軸索末端は、ニューロンの本体とその樹状突起に隣接しています。 それらの接続の場所はシナプスと呼ばれます。 シナプス相互作用により、ニューロンは着信信号に応答し、特定の状況下で、スパイクと呼ばれる独自のインパルスを生成できます。
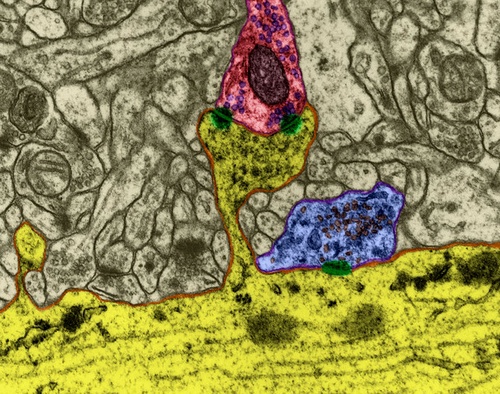
神経伝達物質の割り当てにより、シナプスでの信号伝達が発生します。 軸索に沿った神経インパルスがシナプス前終末に入ると、このシナプスに特徴的な神経伝達物質分子がシナプス小胞から放出されます。 信号を受け取るニューロンの膜には、神経伝達物質と相互作用する受容体が位置しています。
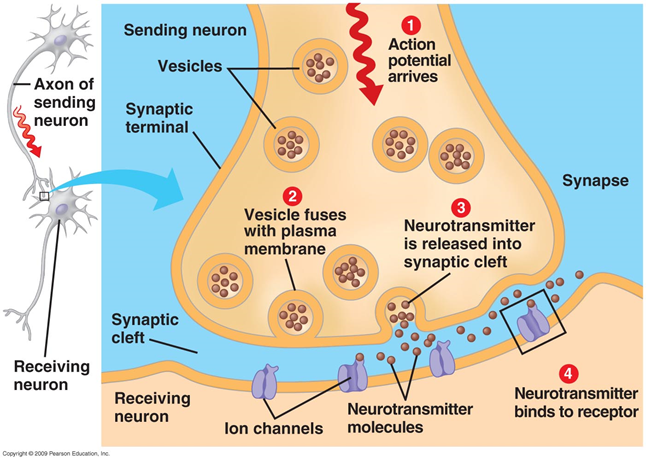
図1.化学シナプス
シナプス間隙に位置する受容体は、ほとんどがイオンチャネル型です。 つまり、それらはニューロンの膜を介してイオンを輸送できるイオンチャネルです。 神経伝達物質は受容体に作用するため、イオンチャネルが開きます。 その結果、どのチャネルが影響を受け、それに応じてシナプスのタイプに応じて、膜は脱分極または過分極されます。 興奮性シナプスでは、チャネルが開き、主に細胞に陽イオンを通過させ、膜が脱分極します。 抑制性シナプスでは、細胞から陽イオンを除去するチャネルが開き、膜の過分極につながります。
特定の状況では、シナプスはその感受性を変化させる可能性があり、これはシナプス可塑性と呼ばれます。 これにより、一部のシナプスがより多くなり、他のシナプスが外部信号の影響を受けにくくなるという事実につながります。
同時に、多くの信号がニューロンのシナプスに到着します。 抑制性シナプスは、膜電位をケージ内の電荷の蓄積に向けてシフトします。 それどころか、シナプスの活性化はニューロンを放電させようとします。 総脱分極が開始しきい値を超えると、活動電位またはスパイクと呼ばれる放電が発生します。
神経伝達物質の放出後、特別なメカニズムがそれらの利用と再捕捉を確実にし、シナプス間隙と周囲のシナプス空間のクリアにつながります。 スパイクに続く不応期の間、ニューロンは新しいインパルスを生成できません。 この期間の持続時間は、ニューロンが可能な最大生成頻度を決定します。
次に、あまり知られていない事実について説明します。
軸索に沿って伝播する活動電位が受容者のニューロンに到達すると、神経伝達物質がシナプス間隙に放出されます。 これらのメディエーターは、シグナルを受け取るニューロンの膜電位の全体的な変化に対するシナプスの寄与を決定します。 しかし、いくつかのメディエーターはシナプス間隙の外側に落ち、ニューロンとそれらを囲むグリア細胞によって形成された空間に広がります。 この現象は、スピルオーバーと呼ばれます(スピルオーバー(英語)-オーバーフロー、オーバーフロー)(Kullmann、2000)。 さらに、神経伝達物質は非シナプス軸索終末およびグリア細胞から放出されます(図2)。 シナプス外の神経伝達物質の濃度は、シナプスの裂け目よりもはるかに低いです。 しかし、これらの「広がる」神経伝達物質には、多くの興味深いことが隠されています。
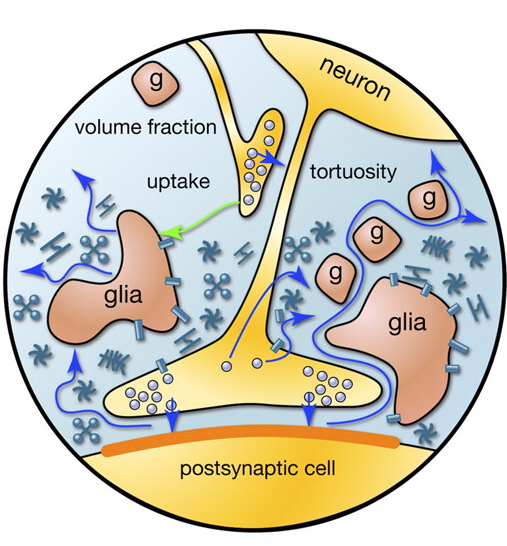
図2.シナプス間隙の外側のメディエーターのソース(Sykova E.、Mazel T.、Vagrova L.、Vorisek I.、Prokopova-Kubinova S.、2000)
シナプスの境界を越えて神経伝達物質を投げるソースの数と構造を推定してみましょう。 このために、下表(表1)にある皮質のパラメーターの定量的推定値を使用します(Braitenberg V.、Schuz A.、1998)。
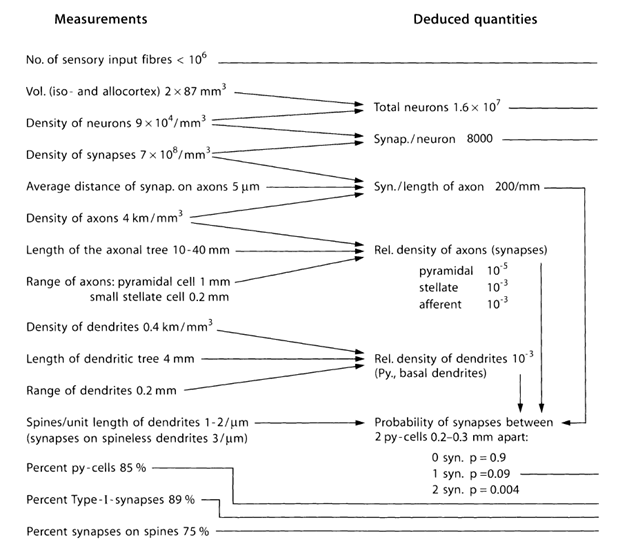
表1.マウス脳の研究で得られたパラメーターの要約表(py-cell-錐体細胞、I型-2つの錐体細胞間のシナプス)(Braitenberg V.、Schuz A.、1998)
シナプスの大部分(90〜95%)はニューロンの本体ではなく、その樹状突起上にあることを思い出させてください。 樹状突起は、ニューロンのいわゆる樹状ツリーを形成する細い分岐プロセスです。 以下の図では、樹状の木は黒で強調表示され、軸索は灰色で強調表示されています。 さまざまなタイプのニューロンの場合、樹状ツリーの形式は異なりますが、一般的な原則は変わりません。樹状ツリーは多くの分岐プロセスで構成され、ニューロンのシナプス結合の最大密度は小さな空間領域に属します。 主なタイプのニューロンの場合、約200μmです(図3、図4)。

図3.星型ニューロンの構造、ルーラー-0.1 mm(Braitenberg、1978)
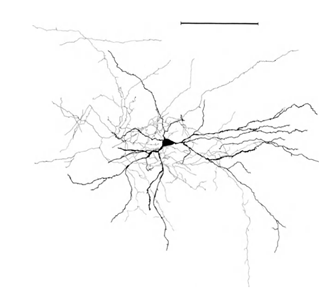
図4.錐体ニューロン、ルーラーの構造-0.1 mm(Braitenberg、1978)
ニューロンの分岐軸索は、他のニューロンの樹状突起との接触(シナプス)を形成します。 樹状突起上のシナプス間の平均距離は0.5マイクロメートルです。 軸索上のシナプス間の平均距離は5マイクロメートル、つまり10倍です。 驚くことではないが、軸索は樹状突起よりも約10倍長い。
ほとんど(75%)のシナプスは樹状突起棘上にあり、これは錐体細胞に最も特徴的です(図5)。
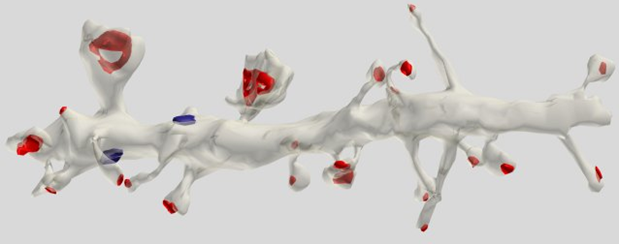
図5.錐体細胞の樹状突起セグメント。 赤色のシナプスは棘に、青色は樹状突起の幹にマークされています(クリステン・M・ハリス博士)
実際の解剖学的および生理学的データに基づくコンピューターシミュレーションでは、たとえば、グルタミン酸が、隣接するシナプス間の距離(0.5μm)に相当する半径でNMDA受容体を活性化するのに十分な量でシナプス間隙を超えて広がることができることが示されました(Rusakov DA、カルマンDM、1998年)。 スピローバー後の神経伝達物質のかなりの濃度が約1〜2μm長の樹状突起の領域で観察されると仮定することができます。 そのようなサイトでは、この樹状突起に属する2〜4個のシナプスを見つけることができます。
長さ5μmの樹状突起の部分を撮影すると(図6)、その上にあるシナプスの予想数は約10になります。
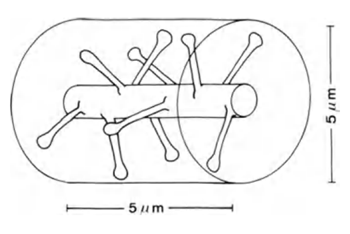
図6.樹状突起のプロット(Braitenberg V.、Schuz A.、1998)
しかし、一部のニューロンの樹状分岐は、他のニューロンの分岐と密接に絡み合っています。 それらはすぐ近くで互いに通過します。 樹状突起スパインの高さにより、1つの樹状突起に属するシナプスは、それ自身のシナプスよりも別の樹状突起の表面に近い場合があります。
シナプスが皮質空間に均等に分布している場合、約100個のシナプスが、高さ5μm、直径5μmの円柱状の体積(上記の図)に落ち、シナプスの分布密度は7x10 8 / mm 3になります。 つまり、ブランチ自体に直接配置されているものの10倍です。 実際、脳の容積のかなりの部分がグリア細胞とニューロンの体で占められており、シナプスの充填密度がさらに増加しています。 ただし、ニューロンはさまざまな神経伝達物質と連携するため、これも考慮する必要があります。
ここで、シナプス外メディエーターの密度分布の観点から、このような結合構造の意味を理解してみましょう。 このために、単純化されたモデルを使用します。 ニューロンを囲む条件付きボリュームを取得し、ニューロンに番号を付けます。 これらの各ニューロンには次のものがあります。
•選択したニューロンの樹状突起とのいくつかのシナプス接触。
•いくつかの「適合」、つまり、他のニューロンとのシナプスが選択されたニューロンの樹状突起に近接して配置される場所。
樹状ツリーを、条件付きソースが均等に分散された1つの長いブランチとして想像してみましょう(図7)。 このブランチの各ソースに対して、それを担当する周囲の空間からのニューロンの数を指定できます。 周囲のニューロンのそれぞれには、樹状突起上にランダムに分布する複数のソース接点が一度にあります。 要素d iを持つベクトルDによるこの相関を示します。
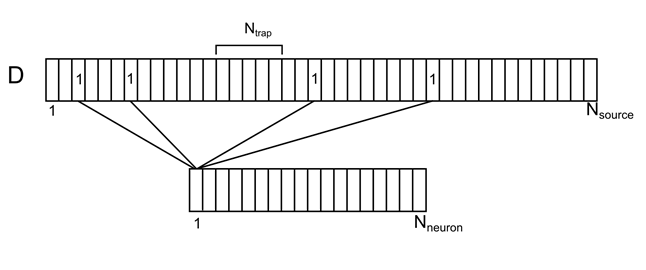
図7.周囲のニューロンと樹状突起上のそれらの接触の相関
次の表記法を紹介します。
N ニューロン -環境ニューロンの数
N source -1つのニューロンのソースの数
N トラップ -神経伝達物質の密度のレベルを作成するソースの数(シナプストラップ)
今、環境からのいくつかのニューロンがスパイクを与えたと仮定します。 これは、ニューロンの観察に利用できる信号として認識できます。 N sigを示します -情報信号を作成するアクティブなニューロンの数。 この信号をバイナリベクトルSで書き込みます。
最も極端なものを除く樹状突起上のすべての位置について、式に従ってメディエーター密度を考慮します
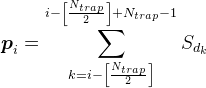
たとえば、下図に示す信号の場合、マークされたシナプストラップの密度は2(1番目と4番目のニューロンからの信号の合計)になります。
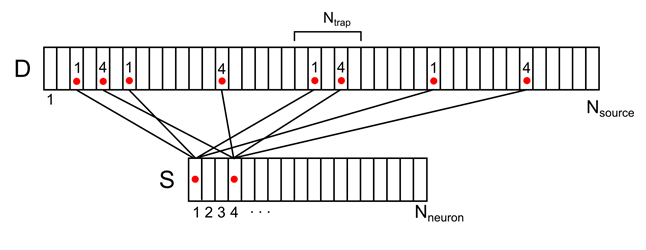
図8. 2つの環境ニューロンの活動の樹状ツリーへのマッピング(接続と番号付けの一部のみを示しています)
任意の信号について、樹状突起上のメディエーターの密度パターンを計算できます。 この密度は0からN trapの範囲です。 シナプストラップを形成するすべてのソースがアクティブな場合、最大値が達成されます。
実際のラット皮質に特徴的なパラメーターの平均値を使用します(Braitenberg V.、Schuz A.、1998)。 それらに基づいて、次のモデルパラメータを取得します。
N ニューロン = 650
N ソース = 25000
N トラップ = 15
信号は、たとえば皮質ニューロンの1.5%などの活動によってエンコードされていると仮定します。
N sig = 10
N sigユニットで構成される任意の信号に対して、メディエーター密度が正確にKである樹状突起に少なくとも1つの場所がある確率を計算するのは簡単です。特定のパラメーターについて、確率は次の値を取ります(表2)。
| K | P |
| 0 | 0.984 |
| 1 | 1 |
| 2 | 1 |
| 3 | 0,996 |
| 4 | 0.287 |
| 5 | 0.016 |
| 6 | 0.001 |
| 7 | 0 |
| 8 | 0 |
| 9 | 0 |
| 10 | 0 |
| ... |
表2.特定の密度の少なくとも1つのトラップを見つけるための確率表。 最初の列は、トラップ内のアクティブなソースの必要数です。 2つ目は、樹状突起上の少なくとも1つの場所を見つける確率です。
つまり、実際の皮質の構成に近いパラメーター値、ニューロンの約1.5%に影響するボリューム信号の場合、次のことが当てはまります。
•シグナル軸索の50%が交差する樹状突起にトラップがあるニューロンの約1.6%があります。
•ほぼすべてのニューロンには、信号軸索の少なくとも30%が交差するトラップがあります。
この結果の意味は非常に興味深いです。 皮質内の情報が、比較的少数(N sig )のコンパクトに配置されたニューロンの同期アクティビティによって何らかの形でエンコードされているとします。 これは、脳のすべての活動に関するものではなく、ニューロンが1からN個のニューロンに番号付けされている小さなボリュームでの情報処理に関するものです。 コードの組み合わせSの数が制限され、N dictの容量を持つ特定の辞書Tを形成するとします。 同じ場所が一度に2つの信号に「応答」する確率を計算することができます。 以下の表に、10,000個の信号のディクショナリに対するこのような計算の結果を示します。
| K | P エラー |
| 3 | 0.00399 |
| 4 | 1,05E-05 |
| 5 | 1.89E-08 |
| 6 | 2,33E-11 |
| 7 | 0 |
| ... |
表3.メディエーター密度のさまざまなレベルでのシナプストラップの一意性違反の確率表
K = 3の場合、トラップは特定の選択性を持ちますが、エラーに対する保証はありませんが、すでにK = 5の場合、特定の空間パターンにかなり明確に対応し始めます。 これは無限のセットからの任意の信号には当てはまらないことを思い出してください。しかし、十分に大きいとはいえ、アクティビティの離散的に許可された状態のセットがある場合。
つまり、脳に特徴的な軸索および樹状突起の構造の意味は、各ニューロンの樹状突起表面、つまり神経伝達物質のあらゆる種類の組み合わせに対応する場所に、トラップの豊かな空間を作り出すことであることがわかります。 トラップ内の神経伝達物質の密度により、空間的に分布した信号を判断できます。これは、近隣の多数のニューロンの同期活動の合計です。
そして、いわゆる代謝型受容体はニューロンの表面にあり、神経伝達物質の濃度が低い場合でも単一のニューロンスパイクを引き起こす可能性があるため、これらのスパイクは自発的と呼ばれ、これらとは異なる膨大な数の異なる信号に応答できることがわかりますニューロンのシナプスを調整できます。
実際、今ではこの記事が始まった樹状突起棘の目的を説明できます。 理論的には、すべてのシナプスは樹状突起またはニューロンの本体に直接配置できます。 これは、シナプスの直接の働きと、誘発された活動の出現に寄与するシナプスの能力には影響しません。 しかし今では、樹状突起スパインの指定は、異なるニューロンのシナプスが「混合」された空間構造の作成であり、表面だけでなく周囲の樹状突起にも影響を及ぼすスピローバーとの能力を獲得するものと想定できます。 そのような「原則的にささいなこと」が得られます。
元の記事は、「波の歴史」自体がこれからどのように生まれるかを示していますが、これは別の会話です。
結論として、少し自己利益があります。 ロシア語のオリジナル記事。 Dmitry Shabanovによる英語への翻訳があります(彼は非常に尊敬しています)が、彼(翻訳)は完全にはほど遠いです。 誰かが間違いを調べて指摘する機会があれば、私は非常に感謝します。 参照による文書は、コメントのためにすべての人に公開されています。
元の記事への参照
1.(2014)。 Human Connectomeプロジェクトから取得: www.humanconnectomeproject.org
2.(2014)。 ALLEN Mouse Brain Connectivity Atlasから取得: connectivity.brain-map.org
3.ブルーム、BH(1970)。 許容エラーを伴うハッシュコーディングのスペース/時間のトレードオフ。 ACM T. 13(7)、422-426の通信。
4. Braitenberg V.、Schuz A.(1998)。 皮質:神経接続の統計と幾何学、第2版。
5. Braitenberg、V.(1978)。 皮質の建築学:一般的および面積的。 MAB BrazierおよびH. Petsche(eds)、大脳皮質のアーキテクトニクス(pp。443–465)。 ニューヨーク:Raven Press。
6. Coster、H.(1975)。 電気機械的応力と膜構造に対するpHの影響。 Biochim Biophys Acta 13; 382(2)、142-146。
7.博士 クリステン・M・ハリス。 (bd)。 シナプスWeb。 synapses.clm.utexas.eduから取得。
8.福島健一(1980)。 ネオコグニトロン位置の変化の影響を受けないパターン認識のメカニズムの自己組織化ニューラルネットワークモデル。 生物サイバネティックス、36(4)、193-202。
9.グロスバーグ、S。(1987)。 競争学習:インタラクティブな活性化から適応的共鳴まで。 認知科学N11、23-63。
10. Hebb、D.(1949)。 行動の組織。 ニューヨーク:ジョン・ワイリー&サンズ。
11.ホジキン、A。a。 (1952)。 膜電流の定量的記述と神経の伝導と興奮へのその応用... J. Physio l。 117、500-544。
12.イジケビッチ、EM(2007)。 神経科学の動的システム:興奮性とバーストの幾何学。 ロンドン:MIT Press。
13. Kullmann、DM(2000)。 哺乳類の脳におけるグルタミン酸とGABAによって媒介されるスピルオーバーとシナプスのクロストーク。 Prog Brain Res、125、339-351。
14.クラモトY.(1984)。 化学振動、波、および乱流。 ドーバー出版。
15. Liibke J.、Markram H.、Frotscher M.、Sakmann B.(1996)。 発達中のラット新皮質の第5層錐体ニューロンによって確立されたオータプの頻度と樹状突起の分布:同じクラスの隣接ニューロンのシナプス神経支配との比較。 Neurosci 16(10)、3209-3218。
16.マレンカRC、ニコルRA(1999)。 長期増強-10年の進歩? Science 285(5435)、1870-1874。
17.マイケル・T・リッパート、高垣健太郎、徐維if、黄小英、建永W。 (2007)。 ラットの皮質活動の電圧感受性色素イメージングの方法。 J Neurophysiol 98、502-512。
18. Pitts W.、McCulloch WS(1947)。 私たちが普遍性をどのように知っているか:聴覚および視覚形態の知覚。 ブル。 数学。 Biophys V. 9、127-147。
19.ローゼンブラット、F。(1962)。 神経力学の原理:パーセプトロンと脳メカニズムの理論。
20. Rusakov DA、Kullmann DM(1998)。 海馬におけるシナプス外グルタミン酸の拡散:超微細構造の制約、取り込み、および受容体の活性化。 Neurosci 18(9)、3158-3170。
21. Sheng、M.、Nakagawa、T.(2002)。 神経生物学:移動中のグルタミン酸受容体。 Nature、417(6889)、601-602。
22. Sheng、M.、Sala C.(2001)。 PDZドメインと超分子複合体の組織。 Annu Rev Neurosci、24、1-29。
23. Sykova E.、Mazel T.、Vagrova L.、Vorisek I.、Prokopova-Kubinova S.(2000)。 細胞外空間拡散および病理学的状態。 脳研究の進展、155-178。
24. Tovar KR、Westbrook GL(2002)。 海馬シナプスの可動NMDA受容体。 ニューロン、34(2)、255-264。
25. W.-F. 徐、X.-Y。 Huang、K。Takagaki、およびJ.-Y. ウー (2007)。 視覚的に誘発された皮質波の圧縮と反射。 ニューロン、55、119-129。
26. Y. LeCun、Y。Bengio。 (1995)。 画像、音声、時系列のたたみ込みネットワーク、マサチューセッツ州アービブ、編集者、脳理論とニューラルネットワークのハンドブック。 MIT Press。
27. J.P. シュヌローヴァ、Z.M。 グボズディコバ。 (1971)。 皮質の感覚運動領域のニューロンの直接電気刺激に対する反応。 コレクション「大脳皮質の神経活動の組織化に関する研究」(p。158-180)。 モスクワ:科学。
28. Nicholls J.、Martin R.、Wallas B.、Fuchs P.(2003)。 NeroからBrain(第4版)。
29. Pribram、K.(1971)。 .
30. , . . (2007). . .
31. , . (2014). . aboutbrain.ru: www.aboutbrain.ru/programs
32. , . (2014). . aboutbrain.ru: www.aboutbrain.ru/programs