私たちは、神経生物学の基礎とニューラルネットワークの理論の改定から、提案されたモデルに含まれる新しいものへと移行しなければならない瞬間に来ました。 サイクルを読み始めたばかりの人のために、 最初のパートから始めることをお勧めします。
実ニューロンの動作の説明に戻りましょう。 一部のニューロンからの軸索を介した信号は、他のニューロンの入力に到着します。 化学シナプスでは、神経伝達物質が放出されます。これは、シナプスのタイプに応じて、信号を受信するニューロンに活性化または抑制効果をもたらします。 シナプスの感度は変化しますが、全体の興奮に対するこのシナプスの寄与を決定します。 合計の影響が特定のしきい値を超えると、膜は脱分極し、ニューロンはスパイクを生成します。 スパイクは単一のインパルスであり、持続時間と振幅は、どのシナプスアクティビティがそれを生成したかに依存しません。
ニューロンのインパルスアクティビティに触発された最も単純なモデルは、しきい値加算モデルです。 さらに、スパイクをバイナリ信号と比較できるという事実に基づいて、加算器の入力と出力は0と1のみの値を取ると考えられています。そのスケールの設定は、ビートごとに応答信号を提供するか、サイレントにする必要があります。 これは非常に論理的です-一定の入力画像は、出力での一定の結果に対応します。
しきい値加算モデルを現実に近づけようとする場合、最初に行う必要があるのは、入力アクティビティの画像が厳密に同期していない可能性があると想定することです。 つまり、異なる入力の信号は、それぞれの周波数を持つパルスでエンコードできます。 この仮定では、入力の状態のインスタントピクチャを使用することはできなくなりました。 指標となる時間間隔を選択し、この間隔で蓄積されたアクティビティパターンを使用する必要があります。 入力信号の異なる周波数では、ある間隔で、入力パルスはニューロンを活性化するのに十分なクラスターを形成し、他の場合は真空を与えてニューロンを非活性化したままにします。 したがって、ニューロンの応答は、入力信号の周波数と対応するシナプスの感度に依存する独自の周波数を取得します。
このような推論は、線形加算器としてのニューロンのモデルにつながります。ニューロンの入力での信号のレベルとその応答は、2つのレベルではなく、スパイク繰り返し率に対応するスカラー量によって記述されます。 線形加算器への移行により、シミュレーションを大幅に簡素化し、神経活動の初期の化学的性質を部分的に忘れることができます。
しかし、すべてには始まりがあります。 情報が脳に入るためには、外の世界と相互作用するニューロンが必要です。 この相互作用の感度は、シナプスの感度とは大きく異なります。 単一のニューロンの数が数万個であるシナプスでは、一定の容量を持つ小胞から神経伝達物質が放出されます。 メディエーターの最小放出量子は、約7,000分子の一部です。 感覚ニューロンは完全に異なるボリュームで動作します。 したがって、視神経スティックは文字通り2つの光量子から活性化され、嗅覚系受容体ニューロンはわずかな臭気分子のみを検出できます。 このような高い感受性は、内部信号増幅メカニズムによって達成されます。
ニューロンと環境との相互作用は、タンパク質分子-細胞内の状態を変化させる外部化学効果を可能にする受容体により発生します。 特定のタイプの受容体と相互作用する物質は、リガンドと呼ばれます。 シナプス受容体の場合、リガンドはニューロンの相互作用を提供するまさに神経伝達物質です。
隣接する受容体が結合して二量体を生成し(下図)、結合すると受容クラスターを形成します。
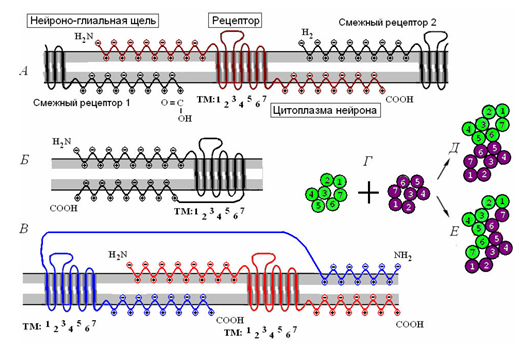
受容体のクラスター化。 Aは単一の受容体であり、周囲の受容体との相互作用です。 Bは単量体受容分子です。 Bは受容二量体です。 G-接触している2つのモノマー(D)とラマン(E)ダイマーの組み合わせ。 (Radchenko、2007)
受容体はイオンチャネル型と代謝チャネル型に分類されます。 イオンチャネル型受容体は、膜を通して荷電イオンを輸送するイオンチャネルを作成し、膜電位を変化させます。 リガンドがイオンチャネル型受容体と相互作用すると、リガンドはイオンチャネルの導電性を変化させ、イオンチャネルを開閉します。 これらの受容体はシナプス間隙に位置しています。 彼らの共同作業により、外部信号の追加によってスパイクが発生するかどうかが決まります。
代謝型受容体は性質が多少異なります。 それらはシナプスの外側にあり、イオンチャネルを作成しません。 リガンドと相互作用すると、細胞内メディエーターの働きを開始し、初期シグナルを数十万回増幅し、最終的にニューロンを活性化します。 これは、代謝型受容体のリガンドである物質のごく一部であっても、対応するニューロンにスパイクを引き起こす可能性があることを意味します。 このようなメカニズムは感覚ニューロンによって使用され、それらが重要でないサードパーティの影響に応答できるようにするのは彼です。 しかし、知覚とは別に、代謝型受容体は、一般的に脳の活動のかなりの部分を担っています。
ニューロンの特徴的な刺激は、そのような入力信号の画像であり、シナプスの感度の画像と一致します。 入力信号が特徴的な刺激に近いほど、ニューロンによって生成されるスパイクの頻度が高くなります。 この反応は誘発活動と呼ばれます。 それは何らかの形で現在の画像に応答したすべてのニューロンで発生します。 しかし、誘発された活動は、脳全体の活動のほんの一部です。 主なアクティビティは、いわゆるバックグラウンドアクティビティに分類されます(下図)。

刺激および背景(自発的)活動に対するニューロンの反応
バックグラウンドアクティビティは、時々発生する単一のスパイクで構成されます。 このような癒着は、誘発された活動の有無に関係なく現れるため、自然発生と呼ばれます。 外部情報から脳を保護すると、感覚ゾーンの自発的な活動は弱まることはなく、むしろ増加するだけです。 単一のニューロンの場合、その自発的な活動は一連のランダムなスパイクのように見えます。 しかし、皮質の1つのゾーンのニューロンの場合、この活動は共通のリズムに発展し、全体として大きな電気的振動を生み出します。 これらの振動は、実際に脳波検査と呼ばれる、頭皮に電極を適用することによって修正することができます。
皮質のさまざまなゾーンと人間のさまざまな状態では、そのようなリズムの頻度とレベルが特徴的です。 皮質の別のゾーンからのリズムは、このゾーンの真上に位置する皮膚の領域で最も強くトレースされます。 したがって、頭の表面に均一に分布した電極のセットを使用して脳のリズムを記録します。 結果は一連のグラフのように見え、それぞれが独自の電極から記録されます(下図)。

脳波の例。 振幅の急激な増加は、てんかん発作の開始に対応します
誘発された活動がイオンチャネル型受容体の作用の結果である場合、自発的な活動は代謝型受容体の活性化の結果です。 軸索に沿って伝播する規則的または自発的な活動の衝動が受容者のニューロンに到達すると、それらはシナプス間隙へのメディエーターの放出を引き起こす。 このメディエーターは、誘発ニューロン活動のプロセスへのシナプスの寄与を決定します。 その後、再キャプチャされ、シナプスが元の状態に復元されます。 しかし、メディエーターの一部はシナプスの裂け目を越えて排出され、ニューロンの体とそれらを取り巻くグリア細胞の体によって形成された空間に広がります。 この現象は、英語のスピルオーバーからのスピルオーバーと呼ばれます-オーバーフロー、オーバーフロー。 近くのシナプスは、いわゆるシナプストラップを形成します。 これらのトラップでは、抑制および活性化メディエーターが干渉パターンを作成します。 これは、インパルスの各組み合わせが、膜内の特定のユニークな場所にメディエーター波腹を引き起こすことを意味します。 A.N. ラドチェンコは、そのような腹の場所にある代謝型受容体クラスターがその特性を変化させ、その後、同じ周囲のインパルスパターンの繰り返しに応答できることを示しました(Radchenko、2007)

代謝型受容クラスターのさまざまな状態(Radchenko、2007)
ラドチェンコは、代謝型受容体クラスターに特徴的ないくつかの状態を説明し、これらの状態をさまざまな記憶段階と比較しました(上記の図)。 細胞の状態に影響を与える受容体の能力は、それらの分岐末端の位置に依存します。 それらの間の距離が大きければ大きいほど、外側の端が表面上に突き出し、内側の端が細胞環境に浸透します。 したがって、距離が大きいほど、受容体の感度が高くなります。 距離が短くなると、受容体の端が膜に沈み、受容体はニューロンの状態に影響を与える能力を失います。 距離は、ニューロンの膜電位によって制御できます。 脱分極は膜の端部をそれぞれ細胞の外側と内側に押し出し、過分極はそれらを反時計回りに膜に引き込みます。
最初に、代謝型受容クラスターにはヒステリシス特性があります。 ヒステリシスは、システムの動作がその背景によって決定されることを意味します(下のグラフ)。 一定の安定した飽和状態に移行すると、システムは労力を取り除いても復帰しませんが、この位置に留まります。 この位置から取り外すには、特定のしきい値を超えた後、システムを同じ「固着」特性を持つ別の安定状態に移行させる逆方向の力を加える必要があります。
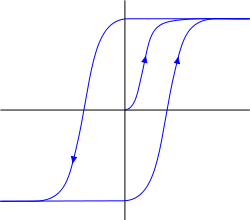
ヒステリシス
ヒステリシスは、受容体にトリガーの特性を与えます。 受容体は、メディエーターの腹に反応して、膜からの放出状態と膜への浸漬状態の両方で「付着」することができます。 最初の状態は、ニューロンが周囲の活動の特定の画像に敏感になり、その繰り返しに独自のスパイクで応答できることを示します。2番目の状態は、反対に、ニューロンを免疫にします。 しかし、膜電位の特定の変化によりクラスターが元の状態にリセットされる可能性があるため、この付着は短期的な性質のものです。
Radchenkoは、受容クラスターのこの挙動を短期(短期)記憶と比較しました。 つまり、特定の瞬間に私たちの思考を決定するのは、一方ではニューロンの活動の写真であり、他方では最近のパターンをすばやく復元できるようにするこの活動の痕跡の固定です。 これらのトラックの脆弱性により、メモリ効率が保証されます。 数秒または1分前の考えにすぐに戻ることができますが、数時間または数日が経過するとこの明るさを失います。
メディエーターの影響と膜電位の強い変化により、受容体はヒステリシス特性を失い、クラスターの獲得状態を長時間固定する安定モードになり、長期記憶と比較できます。 さらに、固定位置、押し出された状態、またはdrれた状態に応じて、クラスターは活動の特定の写真に反応するか、その逆になります-それに対して安定した無反応が得られます。 これらの安定した状態は非常に長期間保存することができ、特定の条件下では上書きすることもできますが、永久に保存できます。
さらにこのモデルでは、脳ニューロンには3つの形式の情報ストレージがあるという事実から進みます。 1つの形式は、ニューロンの画像特性を決定するシナプスの感度の変化です。 2番目の形式は、代謝型受容体クラスターの環境活動のパターンの短期トリガー固定です。 3番目は、受容クラスターが安定状態に移行することによる短期的な固定画像の一部である、長期的な固定です。
ニューロンのシナプスの感度は、ニューロンの唯一の特徴的な画像を表します(シナプスの複数のセクションを持つニューロンを考慮しない場合)。 同時に、数十万の代謝型受容クラスターを含むニューロンのシナプス外表面には、この特徴的な画像がどのイベントで現れたかを示す匹敵する数の記憶を保存できます。
ニューロンのインパルスアクティビティは2つのコンポーネントで構成されていることに注意してください。 1つの構成要素は誘発された活動の写真で、どのニューロンが特徴的な刺激を認識するかを示します。 2番目の要素は、条件付きで自発的な活動の写真であり、最初の要素とは比較的独立して存在し、脳の一般的な背景を作成します。 この背景は、誘導された活動ほど強調されていませんが、皮質の空間全体に分布しているため、より広範囲に広がっています。
私は、脳の最大の謎の一つであるバックグラウンド活動であると言うことを恐れていません。 次のパートでは、この謎を解明しようとします。
継続
どこかが短すぎる、理解できない、または不明瞭な場合は、コメントに記入してください。 私の目はすでに非常にぼやけているので、この素材を初めて知覚する人の代わりに自分を置くのは難しいです。 同時に、難しくない場合は、トピックに対する全体的な習熟度を示します。
前のパーツ:
パート1.ニューロン
パート2.要因
パート3.パーセプトロン、畳み込みネットワーク