 私たちに何かが起こると、私たちの脳はそれを修正し、記憶を作ります。 脳で起こる変化は、一般にエングラムまたは記憶の痕跡と呼ばれます。
私たちに何かが起こると、私たちの脳はそれを修正し、記憶を作ります。 脳で起こる変化は、一般にエングラムまたは記憶の痕跡と呼ばれます。
脳を研究する上で、記憶の痕跡がどのように見えるかを理解することが主な問題であることは当然です。 これがなければ、その仕事の生物学的に信頼できるモデルを構築することは不可能です。 記憶の構造を理解することは、脳が情報をどのようにエンコードし、どのように情報を操作するかを理解することに直接関係しています。 これはすべて、今のところ、未解決の謎です。
記憶の局在化に関する研究は、記憶の謎にさらに興味をそそります。 20世紀前半に、カールラシュリーは非常に興味深い実験を設定しました。 まず、彼はラットを訓練して迷路を抜け出す方法を見つけ、次に脳のさまざまな部分を取り除き、同じ迷路に再び走らせました。 そこで彼は、獲得したスキルの記憶に関与する脳の部分を見つけようとしました。 しかし、時々運動能力の重大な違反にもかかわらず、記憶は毎回保存されることが判明しました。 ネズミは常にどこに道を探すべきかを思い出し、頑固に彼を探しました。

これらの実験により、カール・プリブラムは、人気のあるホログラフィック記憶理論を定式化しました。 それによれば、光学ホログラムと同様に、特定の各メモリは皮質のどの場所にも存在せず、その場所のそれぞれに存在するため、皮質の各場所はすべてのメモリを一度に格納します。
かつて、エングラムの検索における非常に高い期待は、シナプス可塑性に関連していました。 シナプスの感度を変更する能力は、これにより記憶のすべてのメカニズムを説明できるという希望を与えました。 シナプスの可塑性の概念は、人工ニューラルネットワークの作成につながりました。 これらのネットワークは、ニューロンが一連の記憶と共通の何かを学習する方法を示しました。 しかし、一般を学ぶことは、別々の記憶を保つこととまったく同じではありません。
神経科学に直接関与していない場合、神経科学者は記憶に関する多くの理論を持っているという印象を持っている可能性が高いですが、どちらが正しいかは確かではありません。 そして、ほとんどの場合、これらの理論は非常に複雑であるため、大衆文学では特に言及されていません。 ですから、それがどんなに驚くべき音であっても、記憶の単一の理論はありません。 つまり、メモリに関連付けられる可能性のあるものについて、さまざまな仮定があります。 しかし、少なくとも何らかの方法でエングラムがどのように見え、どのように機能するかを説明するモデルはありません。
同時に、ニューロンの生物学、記憶の発現、記憶の形成に伴う分子プロセスなどに関する膨大な知識が蓄積されています。 しかし、知識を深めても状況は単純化されず、複雑になります。 研究の主題についてはあまり知られていませんが、空想するのは便利です。 空想の飛行は知識によってあまり制限されません。 しかし、ますます多くの新しい事実が知られると、多くの仮説が自然に消えます。 事実と一致する新しいものを思い付くのはますます難しくなっています。
科学でこのような状況が発生した場合、これは、致命的なエラーの推論の最初の段階のどこかに忍び込んだことの確実な兆候です。 かつて、アリストテレスは運動の法則を策定しました。 彼は彼の前で見たものから進んだ。 アリストテレスは、自然運動と強制運動の2種類の運動があると言いました。 アリストテレスによると、自然な動きは天体のみに固有のものであり、力を加えることなく動くことができるのは天体だけです。 運動のための他のすべての「地上の」体は力の適用を必要とします、さもなければ、動きは遅かれ早かれ中止しなければなりません。 ほぼ2000年の間、これは明らかな真実とみなされていました。 しかし、同時に、何らかの理由で、これらの2千年すべてにおいて、アリストテレスの声明を超える単一の実行可能な理論を構築することができた人はいません。 そして、ガリレオとニュートンがアリストテレスの厄介な間違いを指摘したときのみ、彼らは彼が摩擦の力を忘れていたと言い、私たちに知られている力学の法則を定式化することが可能になりました。 しかし、アインシュタインはそうでしたが、それは別の話です。
神経科学におけるこのような「強制運動」は、今では「祖母のニューロン」であるように思えます。 実際、記憶の理論を構築する際のすべての主な困難は、特定のニューロンと多くの理由で特定のニューロンに厳密に結び付けられるべきではないメモリの検出器の機能を割り当てると、特定のニューロンをリンクすることは非常に難しいタスクであるという事実に関連しています。
次に、ニューロンが「祖母」の傾斜を失った場合にエングラムがどのように探すことができるかを示します。
前のパートでは、均質な要素で構成されるセルオートマトンについて説明しました。 このオートマトンの任意の場所でアクティビティのパターンが作成されると、波面はこの場所から発散します。 この前線の各場所には、この波のみに固有の特定のパターンが生じます。

波がどの場所を通過するときに作成するパターンを覚えている場合、同じ場所で同じパターンを再現し、この場所から新しい波を開始できます。 この新しい波がそのパスを通過するすべての場所で、元の波のパターンが繰り返されます。
有限数の概念で構成される辞書をコンパイルすると、各概念を独自の固有のウェーブに関連付けることができます。 次に、通過する波のパターンによるオートマトンの任意の場所で、この波が伝播する概念を決定することが可能になります。 そして、私たちが必要な波のパターンの断片をこの場所で再現するならば、どんな場所からでも、どんな概念の波でも発射することが可能になるでしょう。
平らなアバターにボリュームを追加できます。

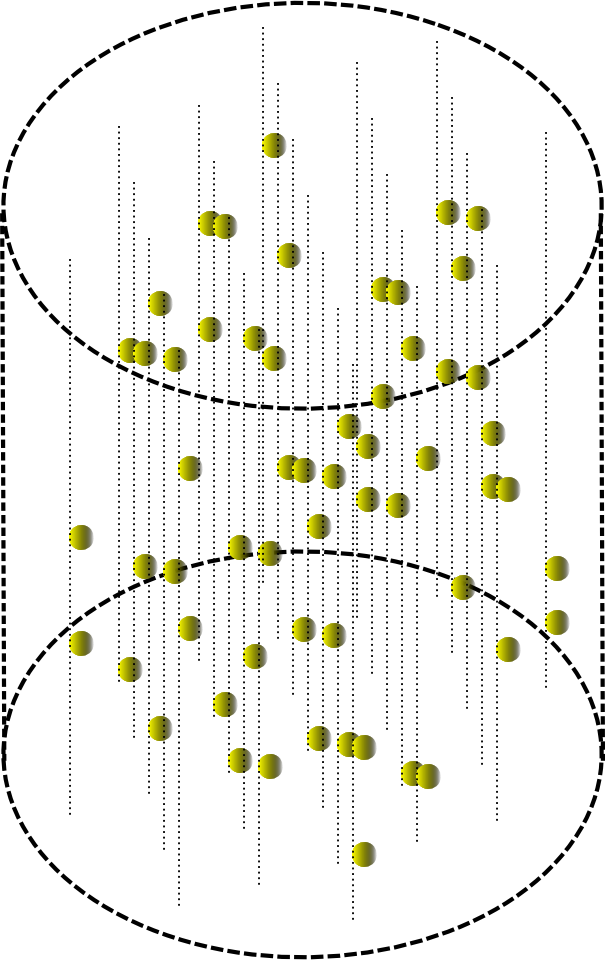
その場合、小さな円筒状のボリューム内の波の通過は、下図に示すようになります。
マシンで情報提供の波を実行する場合、識別子の波を実行します。 これらの波の条件付き「干渉」の写真を思い出すことができます。 これを行うには、情報波が通過した要素であるオートマトンの各場所で、それらを囲む識別子波のパターンを覚えておく必要があります。 この手順により、「キー-値」のペアを覚えることができます。 その後、メモリの識別子の波をオートマトンに起動する場合、マシンの要素はメモリ自体の情報波のパターンを再現します。
キーと値の情報ペアを、マシンの任意の小さな領域で選択的に記憶することも、マシンスペース全体でグローバルに記憶することもできます。 グローバルストレージを使用すると、情報はマシンの領域全体に何度も複製されます。
情報の説明が1つの概念ではなく複数の概念で構成されている場合、これらの概念の情報波を順次拡散させて、そのような説明を自動的に送信することができます。
オートマトンの各固定ボリュームでは、一連の波の通過によりパターンが変化し、それぞれがバイナリベクトルとして記述できます。 概念のシーケンスが説明で重要でない場合、オートマトンの1つの場所で、異なるウェーブによって作成されたバイナリベクトルを論理的にビット単位で結合し、全体の説明ベクトルを取得できます。 十分なビット深度を持つこの合計ベクトルは、それに含まれる概念に関するすべての情報を保持します。
合計ベクトルは容量が大きく、多数のユニットが含まれています。 ハッシュ関数を計算することで、ユニットの数を減らし、ベクトル全体のビット深度を下げることができます。
メモリ識別子は、説明自体と同様に、いくつかの概念で構成できます。 それから彼のために、対応するハッシュを計算することが可能です。 記憶するとき、ソースコードは使用できませんが、ハッシュを受け取りました。
以前に、実際の脳では、ニューロンの樹状ツリーの可能な枝が、セルオートマトン要素の役割の潜在的な候補である可能性があることが示されました。
樹状突起信号のハッシュ関数のニューロン計算
樹枝状の枝は樹状ツリーを形成します(下図)。 ブランチにはペアのブランチのみがあり、閉じたサイクルを形成しません。
リアルニューロンのモデル(EyeWireプロジェクト)
ニューロンのスパイクは、軸索小節の領域で体の膜の脱分極が臨界値に達すると発生します。 体細胞、つまりニューロンの本体の脱分極は、主に樹状分岐からの信号により発生します。 潜在的に、そのような信号は、樹状分岐および樹状突起癒着で生じる電流であり得る。
ナマズへの途中で、分岐部位の異なる枝の信号が相互作用し、ニューロンの体に到達する信号が樹状分岐の信号の一部の機能であることが判明したためです。 ニューロン本体自体の膜電位は、樹状突起ツリーのすべての枝の信号の関数です。 ちなみに、これは形式ニューロンの古典的な考え方に特に反するものではありません。 古典的な形式ニューロンはシナプスでの信号の単純なしきい値加算器であり、樹状分岐の信号のややトリッキーな機能について話していることに注意してください。
説明した解釈では、1つのニューロンのスパイクは、その樹状分岐の信号に対するハッシュ変換のバイナリ結果と安全に呼ばれます。 したがって、ニューロンの活動の全体像は、樹状突起セグメントの活動のハッシュ変換の結果として解釈できると言えます。
ニューロンをアクティブにするには、樹状ツリーで発生するすべてのアクティビティが数ミリ秒の短い時間間隔に収まる必要があります。 このような間隔で、複雑な記述のすべての波が通過した後に生じる樹状突起活動の全体的なパターンが形成されると仮定すると、ニューロンスパイクは、樹状突起セグメントに生じたパターンに関連するハッシュの役割に理想的に適しています。
パターンの干渉を通して記憶するには、識別波と価値の波、つまり保存された情報という2つの波が必要です。 実際の地殻では、これらの波は同時に伝播できます。 同時に、識別子自体はかなり複雑な記述になる場合があります。 皮質では、情報のハッシュと識別子のハッシュを同時に、ただし異なるニューロンによって形成できると想定できます。 原則として、これらのニューロンは異なるタイプのニューロンになります。 最も一般的な皮質ニューロンは、錐体ニューロンと星状ニューロンです。 たとえば、錐体ニューロンの活動が情報のハッシュ関数をエンコードし、メモリ識別子の星形ハッシュ関数のアクティビティをエンコードしていることがわかります。
選択された樹状点
皮質のあらゆる場所で、現在の情報はこの場所にあるニューロンの活動の組み合わせによってエンコードできるという結論に達しました。 それらのスパイクの瞬間的な全体像は、これらのニューロンに対応する樹状突起活動のハッシュ関数として知覚できます。
セルラーオートマトンでは、記憶するために、オートマトンの各要素が十分な長さのハッシュコードの断片を見て覚えることができなければなりませんでした。 識別波は、どの要素を記憶する必要があるかを示し、一連の情報波は、同じハッシュコードが取得された要素のアクティビティの全体像を形成しました。
脳と同様に、これは、各樹状分岐が周囲のニューロンの活動を見て、それを選択的に記憶できるはずであることを意味します。
1つのブランチが1つまたは2つではなく、数千または数百万の異なるニューロン活動の組み合わせを記憶できるようにするには、タスクが非常に興味深いものになります。
ここまでは、主にニューロンの樹状ツリーについて説明しましたが、次に軸索を見てみましょう。 したがって、皮質ニューロンの主な割合は、錐体ニューロンと星状ニューロンに分類されます。 これらのニューロンの軸索は、高度に分岐した側枝によって特徴付けられます。 軸索のシナプス接触の大部分はボリューム上に落ち、その寸法は樹状突起ツリーのサイズに匹敵します(下図)。 この軸索形状により、ニューロンの活動に関する信号が、このニューロンの特定の近傍(半径が50〜70μm程度)にあるこのニューロンおよび他のニューロンのほぼすべての樹状突起枝で利用可能になります。
星ニューロンの構造、定規-0.1 mm(Braitenberg、1978)
信号の可用性は、その近くの各樹状分岐について、ニューロンがこのニューロンの軸索がその近くを通過する場所になるという意味で理解されるべきです。 したがって、ニューロンの活動の瞬間に、スパイクがその軸索に沿って伝播し、神経伝達物質は軸索によって形成されたすべてのシナプスから放出されます。 spiloverによるこれらの神経伝達物質の一部、つまりシナプスを越えた放出は、望ましい樹状分岐に到達することができます。
一般的に、軸索は皮質に沿って、またはそれを超えて広がる可能性があります。 しかし、ほとんどの場合、主要な軸索分岐はニューロン自体を取り巻く空間に落ちます。 樹状突起上のシナプス間の平均距離は0.5μmです。 軸索上のシナプス間の平均距離は5ミクロンです。 樹状突起上の接触の数は、シナプス上の接触の数に等しい。 したがって、軸索の全長は樹状突起の全長の10倍です。 最も近い周囲のニューロン空間は、そのシナプスの約6,000を占めています。 これは、3センチメートルの軸索の長さに相当します。 ここで、これらの3センチメートルが半径10分の1ミリメートル未満の球体に配置され、軸索分岐の性質を理解できると想像してください。 樹状突起の任意のセグメントの近くに隣接するニューロンの軸索が多くあります;さらに、いくつかは、複数回それに接近します。
樹状分岐を直接取り巻くシナプスは、内因性と単純の両方で、近くにあり、この分岐のシナプス外神経伝達物質の源です。 条件付き樹状突起セグメントに沿ったこれらのソースの位置を示しましょう(下図)。 このため、ソースは樹状突起と互換性があり、樹状突起の長さに沿ったおよその位置を観察します。 周囲の樹状ニューロンに番号を付けます。 次に、このブランチの各ソースに対して、このソースを制御する周囲の空間からのニューロンの数を指定できます。 各環境ニューロンは、樹状突起上にランダムに分布する複数のソースを同時に制御できます。 要素D iを持つベクトルDによる樹状分岐上のニューロンとソースの相関を示します。各要素はソースを制御するニューロンの数です。
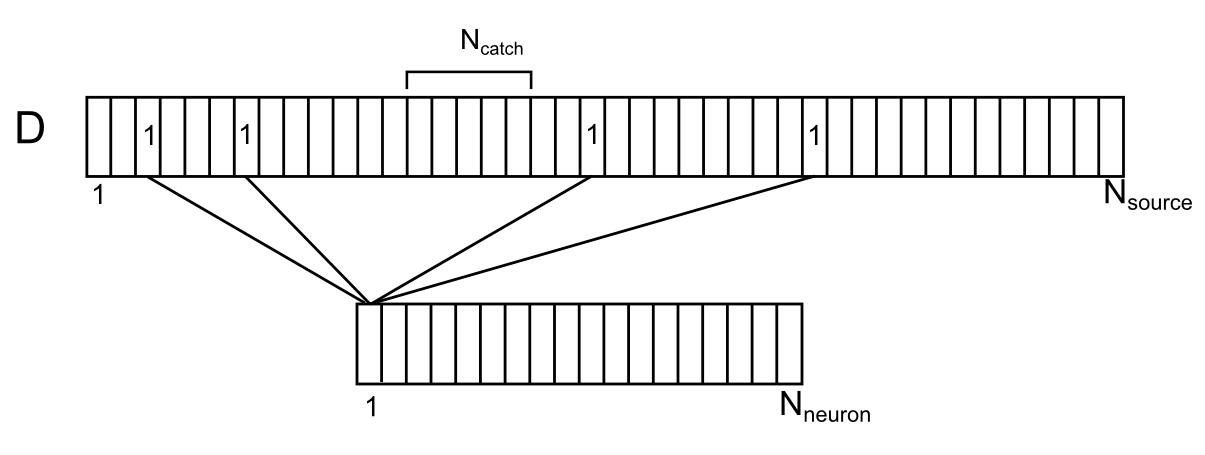
周囲のニューロンと樹状突起上のそれらの接触の相関
N個のニューロンを環境ニューロンの数とし、N個のソースを1つの樹状突起セグメントのソースの数とします。
神経伝達物質がスピラーの後に伸びる距離を自問すると、どのシナプスが樹状突起の選択された位置に影響を与える可能性があるかを判断できます。 Nは、樹状突起の選択された位置に影響を与えることができるソースの数を示します。 これらのソースでは、そのような場所は「トラップ」と呼ばれます。
今、樹状突起の環境からのいくつかのニューロンが癒着を生じたと仮定します。 これは、樹状突起セグメントの観察に利用できる信号として解釈できます。 N sigを示します -情報信号を作成するアクティブなニューロンの数。 この信号を、次元N sourceのバイナリベクトルSで書き込みます 。
最も極端なものを除く、樹状突起上のすべての位置について、式に従って、トラップに入るアクティブなソースの数(信号密度)を決定できます。
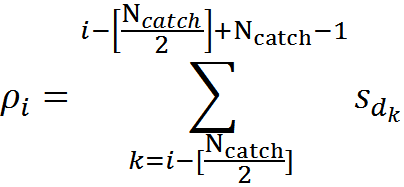
たとえば、下図に示す信号の場合、マークされたシナプストラップの信号密度は2(1番目と4番目のニューロンからの信号の合計)になります。
2つの環境ニューロンの活動の樹状突起セグメントへのマッピング(接続と番号付けの一部のみが示されています)
任意の信号に対して、樹状突起上に作成する密度分布を計算できます。 この密度の範囲は0〜N catchです。 対応するトラップを形成するすべてのソースがアクティブになると、最大値に達します。
実際のラット皮質に特徴的な値を使用し(Braitenberg V.、Schuz A.、1998)、それらに基づいて、モデルの近似パラメーターを選択します。
セクションの長さ= 150μm(300シナプス、シナプス間の平均0.5μm)
樹状突起の高密度分岐の半径= 70μm
皮質のニューロンの密度= 9x10 4 / mm 3
樹状突起に囲まれたニューロンの数(N ニューロン )= 100
樹状突起セグメントのソース数(N source )= 3000
トラップサイズ(N キャッチ )= 15
信号は、たとえばニューロンの10%などのアクティビティによってエンコードされていると仮定します。
N sig = 10
N sigユニットで構成される任意の信号について、樹状突起セグメントに少なくとも1つの場所が存在する確率を計算することができます。信号密度は正確にKユニットになります。 指定されたパラメータの場合、確率は次の値を取ります。
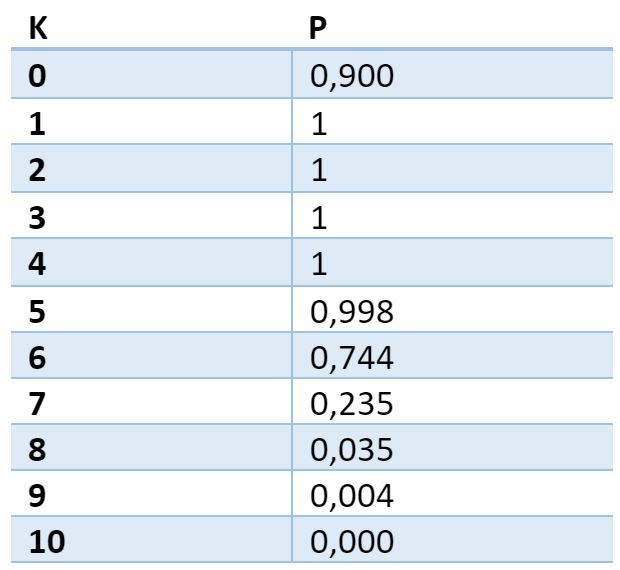
指定された密度の少なくとも1つのトラップを見つけるための確率テーブル。 最初の列は、トラップ内のアクティブなソースの必要数です。 2つ目は、樹状突起上の少なくとも1つの場所を見つける確率です。
この表は、選択されたサラウンド信号の樹状セグメントで1に近い確率で、アクティブなニューロンの少なくとも5つの軸索が収束する場所があることを示しています。 この樹状突起サイトは、選択された信号に関連して選択されたと見なすことができます。 この場所で、どの軸索(シナプス)がアクティブであったかを正確に記憶する場合、これにより、その後、同じシグナルの繰り返しを高精度で検出できます。
検出の精度は、衝突の確率、つまり、選択された場所で軸索が収束する同じニューロンが他のボリューム信号でアクティブになる確率によって決まります。 つまり、たとえば、信号が100個中10個のニューロンの活動によって決定され、これら10個のニューロンのうち5個の組み合わせが選択された場所で記録された場合、衝突については、これら5個のニューロンが他の信号でもアクティブであることが十分です。
トラップに「閉じ込められた」ニューロンの数、つまり、選択された場所で活動が一致したニューロンの数をKで表します。Nsigアクティブニューロンで構成される別の信号が誤認識を引き起こすには、そのニューロンがトラップニューロンと一致する必要があります。 そのようなマッチング信号の数は
 。 可能な信号の総数は
。 可能な信号の総数は 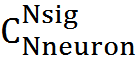 エラー確率
エラー確率
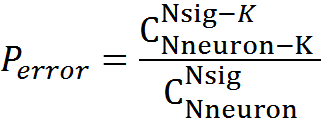
K = 5のモデルの場合、検出エラーの確率は3.34x10 -6になり、K = 6の場合、確率はそれぞれ1.76x10 -7になります。
神経伝達物質の組み合わせによる選択された場所での信号コーディング
各樹状突起部位は、それ自身と隣接する樹状突起の両方のシナプスに囲まれています。 これらのシナプスは、シナプス外神経伝達物質の源です。 樹状突起の選択されたサイトに影響を与えることができるシナプスのそれらは、トラップを形成します。 任意の樹状突起サイトのこのようなシナプスの平均数はN catchです。 Kの値を設定します。これは、サラウンド信号に関連して選択された場所を考慮するためにアクティブにするシナプスの数を決定します。 樹状突起の各場所には、この場所に少なくともK個の交差点を作成する多くの信号があることがわかります。 目的の信号の繰り返しを正確に追跡できるように、この信号に関して選択された場所でK個のソースが機能したという事実を修正するだけでなく、これらが信号に対応するソースであることも確認する必要があります。 つまり、神経伝達物質の放出により、活性化されたシナプスの数だけでなく、今回どのシナプスが機能したかを判断する必要があります。
すでに述べたように、ほとんどのシナプスでは、活動時に1つの「主要な」神経伝達物質が放出され、それに加えて1つ以上の神経ペプチドが放出されます(Lundberg、JM 1996. Pharmacol。Rev. 48:113-178。)(Bondy、CA、et。 1989. Cell。Mol。Neurobiol。9:427-446)。 脳のニューロンに多数の神経伝達物質と神経調節物質が存在するという事実は、そのような多様性の主な機能は、ニューロンの同期活動時の空間の各場所におけるメディエーターと調節物質のユニークな組み合わせの作成であることを示唆しています。 シナプス小胞内の追加の物質は、空間内の各場所で最大の分布多様性を確保するような方法でシナプスに分布していると想定できます。 そうである場合、シナプス活動の特定の組み合わせの検出は、これらのシナプスに対応する放出された物質の一意のセットを決定することになります。
したがって、この信号に特徴的な物質の組み合わせに敏感な検出器が特定の信号に関して選択された樹状突起部位に配置される場合、この検出器の動作は初期信号の繰り返しを示す可能性が非常に高い。
今では、すべての樹状分岐に、周囲のニューロンの信号に関連して選択される場所が常にあることがわかっています。 樹状突起の枝が、これが彼女の選んだ場所が神経伝達物質の特定の組み合わせに反応するはずであることをどのように思い出すことができるかを理解することは残っています。
記憶の要素としてのニューロン受容体
ニューロンの働きを説明すると、周囲の物質がニューロンの受容体を介してニューロンの働きに影響を与えると言いました。 受容体はイオンチャネル型および代謝型です。 イオンチャネル型受容体は、シナプスで放出される神経伝達物質に結合し、その構造の変化を引き起こします。 分子の立体構造は、分子の組成を変えずに空間構造を変化させることです。
イオンチャネル型受容体は、同時にイオンチャネルです。 コンフォメーションの変化により、受容体のイオンチャネルが開き、イオンの動きが生じ、それに応じて膜電位が変化します。
代謝型受容体はイオンチャネルを持たず、異なる働きをします。 ニューロンの内側にある部分は、いわゆるGタンパク質に関連付けられています。 これらの受容体がシグナル伝達物質と相互作用すると、立体構造が変化し、Gタンパク質が放出されます。 これにより、さまざまな結果が生じる可能性があります。 考えられる結果の1つは、ニューロンの局所膜電位を十分にすばやく変化させる、隣接するイオンチャネルのGタンパク質による発見です。 これにより、樹状突起に電流が発生し、樹状突起の付着を引き起こす可能性があります。
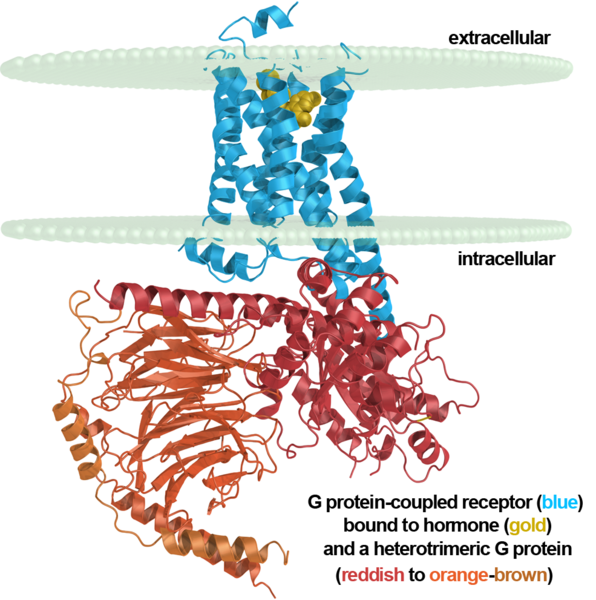
代謝型受容体、ニューロン膜およびGタンパク質
代謝型受容体は主にシナプスの外側に位置し、シナプス外神経伝達物質の特異的な標的です。 受容体は主にクラスター化され、一緒に作用します。 実際、代謝型受容体のクラスターは、神経伝達物質の特定の組み合わせに合わせて調整されたさまざまな検出器です。
代謝型受容体のクラスターは、モデルのエングラム要素の役割に非常に適しています。 数百のそのようなクラスターを各シナプスの隣に配置できます。 それらでは、シナプス外神経伝達物質、すなわち外部環境の隣接シナプスから際立っている神経伝達物質の多くの可能な組み合わせに潜在的に敏感な受容体の組み合わせが事前にランダムに構成されています。
つまり、樹状突起の各部位に多数存在する代謝型受容体は、将来のエングラムの「空白」になる可能性があります。 「空白」からエングラムへの移行は、次のように説明できます。樹状分岐に属する受容体クラスターは、最初は不活性であり、その機能にはまったく影響しないと仮定します。このブランチが暗記を必要とするとき、すべての代謝型受容体にこれを認識させます。そのような信号は、例えば、この分岐の膜のわずかな完全な脱分極であり得る。上で言ったように、このブランチのどこかに確かにお気に入りの場所があります。つまり、このブランチの隣に、一度に複数のアクティブなシナプスがある場所です。枝のこの場所に受容体を持つクラスターがあり、その感度が神経伝達物質のカクテルと一致する場合、このクラスターはアクティブな状態になり、その後常にカクテルの外観に反応する必要があります。これより前に、クラスターが樹状突起枝の機能に影響を与えなかった場合、そのカクテルが現れるときに刺激的なシナプス後電位を作成する必要があります。
代謝型受容体は、このように振る舞う方法を本当に知っていることがわかります。特定の状況では、受容体の外側部分と内側部分が膜に逆らって浸ることがあり、受容体の感度が失われます。または、逆に、受容体の敏感な部分が膜から押し出される可能性があります。この敏感な状態では、受容体はしばらくの間残ることがあり、それから元の状態に戻ることができます-これは短期記憶に対応します。受容体は、そのような敏感な状態を長い間修正できます。適切な条件が形成されると、接着と重合のプロセスが開始され、数日から数週間にわたって受容体が敏感な状態になります。おそらく1か月続く固定プロセスが約1か月間止まらない場合、受容体の状態は永久に、またはより正確には生涯にわたって固定されます。これはすべて、長期記憶の統合のさまざまな段階に対応しています。
代謝型受容体のこの振る舞いを制御するメカニズムは、A。N. Radchenko(脳の情報メカニズム、2007)によって詳細に研究され、説明されています。ところで、ラドチェンコは、コンホメーション転移を伴う代謝型受容体の正確なクラスターがエングラムの要素であることを最初に示唆した。
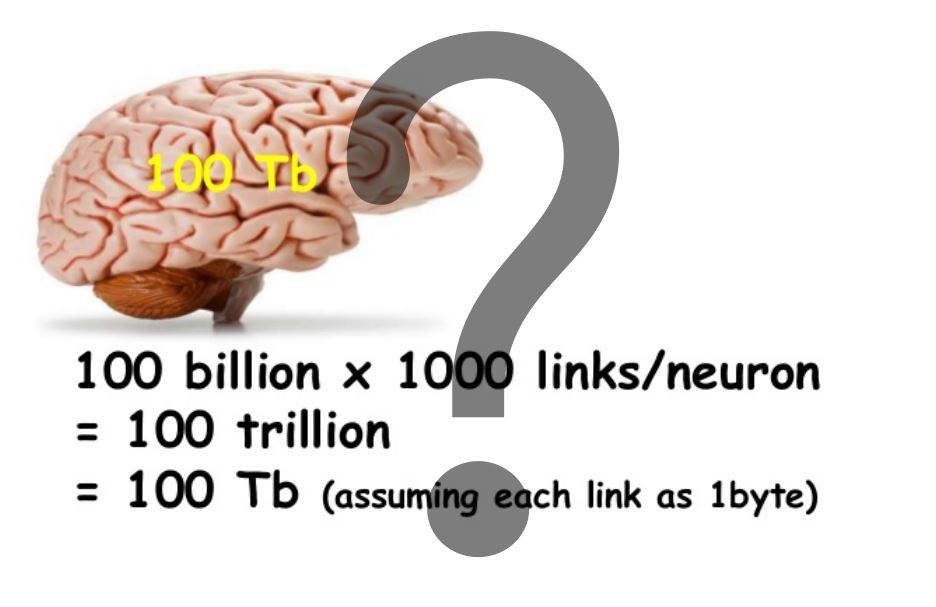
シナプスの可塑性に基づくメモリの場合、容量は非常に単純であると見なされます。このような計算の例をタイトル画像に示します。このモデルでは、樹状突起のメモリ容量は約1000倍大きいことに注意してください。それだけではありません。
軸索と樹状突起の織り合わせによって作成される空間構造では、「選択された場所」のイデオロギーが機能します。つまり、受容体が「ビジネス」であるためには、シナプスが属する樹状突起に関係する必要はありません。樹状突起を介して信号が送信されます。神経伝達物質が細胞間空間に広がるという事実により、単純に幾何学的に近くに配置されている受容体はすべて「作用中」になります。さらに、これは一般に、ニューロンに特異的に属する受容体である必要はありません。
したがって、グリア皮質細胞、プラズマ星状細胞(下図)は、ニューロンと同じ受容体のセットを持っているため、記憶メカニズムの参加者になります。
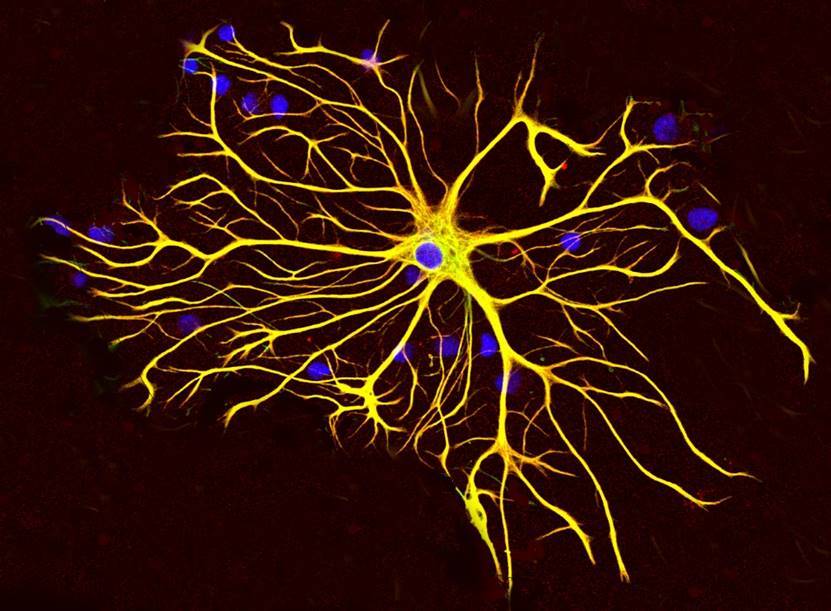
プラズマ星状細胞
皮質では、星状細胞の数はニューロンの数を超えています。皮質の星状細胞には短い分岐プロセスがあります。これらのプロセスにより、それらは近くのシナプスをカバーします(下図)。
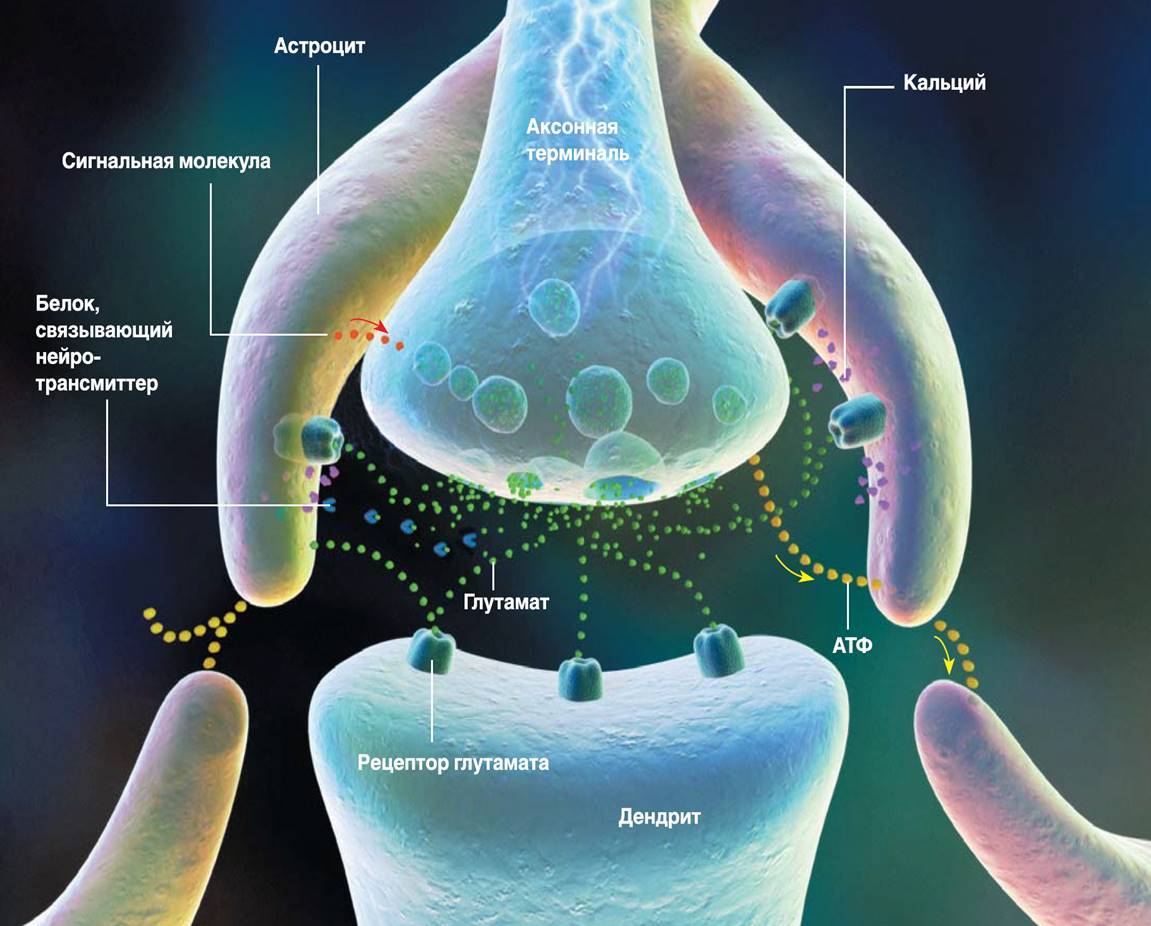
三者シナプス(RD Fields、B。Stevens-Graham、2002)
星状細胞は、対応するメディエーターを排出することによりシナプス応答を増強し、神経伝達物質結合タンパク質の取り込みまたは放出によりシナプス応答を弱めることができます。さらに、アストロサイトは、軸索による神経伝達物質の放出を調節するシグナル伝達分子を分泌することができます。アストロサイトの影響を考慮したニューロン間の信号伝達の概念は、三側シナプスと呼ばれています(RD Fields、B。Stevens-Graham、2002)。三国間のシナプスが、さまざまなメモリシステムの相互作用のメカニズムを実装する主要な要素である可能性があります。
海馬の役割。識別子の情報。リング識別子
記述されたメモリモデルでは、情報画像自体に加えて、メモリが皮質の任意のゾーンに形成されるために、メモリ識別子の信号が送信されます。皮質のゾーンは異なる機能を実行するため、メモリの識別子はゾーンまたはゾーンのグループごとに異なると想定するのが適切です。
一部のよく知られている脳領域は、その特異性により、識別子ソースの役割に適しています。そのため、4倍体の上部結節は、視覚ゾーンの識別子のソースになります。 4重の下部結節は、皮質の聴覚ゾーンの識別子ジェネレーターの役割に適しています。記憶に関連する最も重要な器官は海馬であり、前頭前野の記憶域の重要なジェネレーターの役割に適しています。
1953年、一般にHM(ヘンリー・モレゾン)と呼ばれる患者がてんかんの治癒を試み、両側海馬の除去を受けました(W. Scoviille、B。Milner、1957)。その結果、HMは何も記憶する能力を完全に失いました。彼は手術前に彼と一緒にいたことをすべて覚えていましたが、注意が移るとすぐに何か新しいことが頭から飛び出しました。クリストファー・ノーランの「メメント」映画を見た人は誰でも、会話の内容をよく理解しているでしょう。

Henry Molaison
HMの事例は非常にユニークです。 HMのように完全な両側破壊がなかった海馬の除去に関連する他のケースでは、記憶障害はそれほど顕著ではなかったか、まったくなかった(W. Scoviille、B. Milner、1957)。
海馬の完全な除去は、新しい記憶を形成することを不可能にします。海馬の異常はコルサコフ症候群を引き起こす可能性があり、これはまた、古い記憶を維持しながら現在のイベントを記録できないことを減らします。
海馬の役割についてのかなり一般的な考えは、海馬は現在の記憶の保存場所であり、その後、皮質の空間に再分布されるということです。記述されたモデルでは、海馬の役割は異なります;それは記憶のユニークなキーを作成します。
海馬によって作成されたキーは、投影システムを通じて皮質の対応するゾーンに配布されます。海馬の識別子と情報の説明の干渉が記憶を作成します。この場合、記憶はすぐに「その場で」形成され、海馬と皮質の間を移動しません。この表現は実験データとよく一致しています。確かに、海馬の除去は、記憶の重要なジェネレーターが消えるので、新しい暗記を不可能にします。古い記憶は海馬に依存しなくなったため、そのまま残っています。それらの識別子は、海馬の関与なしに抽出して使用できます。
しかし、提案されている海馬の役割を支持する主な議論は、海馬に見られる機能に関連しており、一見すると記憶機構に直接関連していません。
1971年、ジョンオキーフは海馬の細胞を発見しました(オキーフJ.、ドストロフスキーJ.、1971年)。これらのセルは、内部ナビゲーターのように反応します。ラットが長い廊下に置かれている場合、特定の細胞の活動により、ラットがどこにいるのかを正確に言うことができます。さらに、これらの細胞の反応は、この場所に到達した方法に依存しません。
2005年に、海馬で空間位置をコードするニューロンが見つかり、座標グリッドのようなものを形成しました(Hafting T.、Fyhn M.、Molden S.、Moser MB、Moser EI、2005年)。
2011年、海馬には時間間隔を特定の方法でエンコードする細胞があることが判明しました。彼らの活動は、他に何も起こらなくてもリズミカルなパターンを形成します(Christopher J. MacDonald、Kyle Q. Lepage、Uri T. Eden、Howard Eichenbaum、2011)。
データをキーと値のペアとして保存すると、連想配列が作成されます。連想配列では、キーには二重の機能があります。一方では、1つのペアを別のペアと区別できる一意の識別子であり、他方では、キー自体が検索を非常に容易にする情報を運ぶことができます。たとえば、コンピューターのファイルシステムは連想配列と考えることができます。値はファイルに保存されている情報、キーはファイルに関する情報です。ファイルに関する情報は、保存場所、ファイル名、作成日を示すパスです。写真、追加情報-ジオタグ、写真が撮影された場所の座標。音楽ファイルの場合、アルバム名とアーティスト名。これらのファイルデータはすべて、ファイルを一意に識別するだけでなく、ただし、キーフィールドのいずれかまたはそれらの任意の組み合わせで検索することもできます。キーが詳細であるほど、検索オプションはより柔軟になります。
脳はコンピューターシステムと同じ情報タスクを実行するため、脳によるキーと値のペアの形式でのデータの保存には、検索に最も便利なキーの作成が伴うと想定するのが論理的です。人が扱っている記憶のために、主要な記述子の合理的なセットには以下を含める必要があります。
- 実施場所の表示;
- 空間内の位置の表示。
- イベントの時間の表示。
- 起こっていることの主な意味をアドバイスする一連の概念。記事の内容を説明するキーワードの類似物。
海馬は、場所、空間、時間の位置だけでなく、このデータを正確に使用して、記憶の複雑な情報キーを構成する可能性が非常に高いです。少なくとも、このような多様な機能が1か所に集まった理由は非常によく説明されています。また、記憶の形成に直接関与する場所。
特に興味深いのは、一時的なコーディングです。人間の記憶により、静止画像を思い出すことができるだけでなく、時系列を維持しながら一連のシーンを再現できます。したがって、そのような機会はメモリコーディングシステムに組み込まれるべきです。海馬にはリズミカルなパターンを作成する時間セルがあることが示されています(Christopher J. MacDonald、Kyle Q. Lepage、Uri T. Eden、Howard Eichenbaum、2011年)。パターンの周期的な性質は、海馬が時間を測定してイベント識別子の時間フィールドを作成するために使用するのと同じ手法を使用できることを示唆しています。
時間の経過を説明するには、時計とカレンダーを使用します。どちらもリング識別子に基づいています。 1分は60秒です。これは、60個の識別子が連続して互いに置き換えられ、60秒後に最初の識別子が再び続くことを意味します。同様に、時間あたりの分数、日数時間、月数日、週数日、年数月、世紀数年。つまり、異なる周期性を持ついくつかのリング識別子により、任意の時点を識別できます。

海馬は一時的なリング識別子の同様のシステムを使用しているようで、実際に実験で観察されました。しかし、特に興味深いのは、そのようなシステムにより、記憶を識別するだけでなく、そのシーケンスを再現することも可能になることです。識別子を次々に追跡し、1つのイベントの識別子を取得するためのルールを知っている場合、そのイベントの時系列で後に続くイベントの識別子を簡単に取得できます。
リング識別子は、時間の説明だけでなく、他の多くの状況でも便利であることに注意してください。
このパートで説明したことは、脳の機能を理解するために非常に重要ですが、まだ「魔法」は含まれていません。はい、メモリがどこに、どのような形で保存できるかを示していますが、脳がコンピュータの力を超えたタスクを簡単に解決する方法については何も言いません。もちろん、コンピューターは人よりも速く考えることができますが、人は何が起こっているのかを理解することができます。次の部分では、一般に「感覚」があり、徐々に、ニューロンの働きとホログラフィックメモリのメカニズムを長い間説明しなければならなかった理由が明らかになります。
アレクセイ・レドズボフ
意識の論理。 エントリー
意識の論理。 パート1.セルオートマトンの波
意識の論理。 パート2.樹状波
意識の論理。 パート3.セルオートマトンのホログラフィックメモリ
意識の論理。 パート4。脳記憶の秘密
意識の論理。 パート5.情報分析へのセマンティックアプローチ
意識の論理。 パート6.意味を計算するためのスペースとしての皮質
意識の論理。 パート7.コンテキスト空間の自己組織化
意識の論理。 「指で」説明
意識の論理。 パート8.大脳皮質の空間マップ
意識の論理。 パート9.人工ニューラルネットワークと実際の皮質のミニコラム
意識の論理。 パート10.一般化のタスク
意識の論理。 パート11.視覚および音声情報の自然なコーディング
意識の論理。 パート12.パターンを検索します。 組み合わせ空間