
この一連の記事では、従来のモデルとは大きく異なる脳の波動モデルについて説明します。 参加したばかりの人は最初の部分から読み始めることを強くお勧めします。
海馬が完全に除去されると、HM患者の場合が納得できるように、新しい記憶を形成することができなくなります。海馬の異常はコルサコフ症候群につながり、古い記憶を保存しながら現在の出来事を記録できなくなることもあります。 これはすべて、海馬が記憶メカニズムにおいて重要な役割を果たすことを私たちに確信させます。
海馬の役割に関する伝統的な理論は、脳とコンピューターの類似性に基づいています。 このような考慮事項では、海馬には「ランダムアクセスメモリ」、つまり新しいメモリが蓄積される場所の役割が与えられます。 そして、おそらく夢の中で、これらの記憶は長期記憶の保存に関与する脳の領域に転送されます。 そのような転送のメカニズムは明確ではありませんが、少なくともこれは、海馬の破壊がイベント記憶の形成をブロックする理由を説明するのに役立ちます。
私たちのモデルは、従来のモデルと根本的に異なります。 私たちは、海馬とその後のコピーにメモリバッファリングがありません。 記憶を形成する要素は、適切な場所にすぐに形成されます。 海馬は、皮質の空間に分布する記憶要素を結合する単一の識別子を作成するだけです。 脳の波動モデルは、この識別子が皮質全体にどのように広がるかを説明しています。 そのような識別子の存在は、それによって結合された要素を選択することにより、抽象的な写真ではなく、記憶された出来事の具体的な画像を再現することを可能にします。
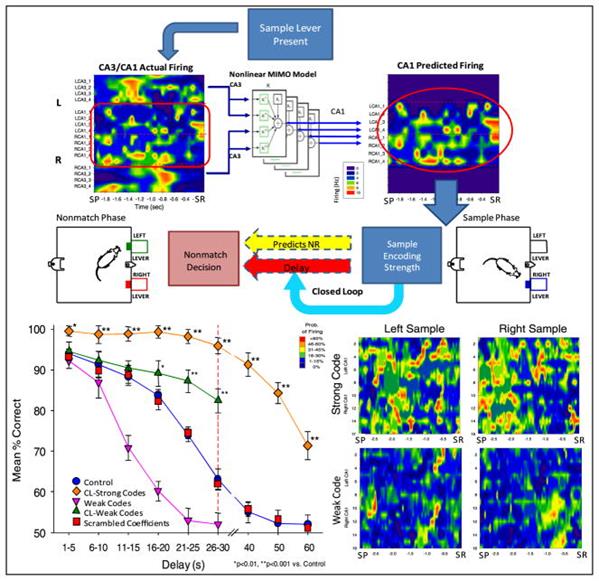
2010年には、ラット海馬の補綴物に関して興味深い実験が行われました(セオドアW.バーガー、ロバートE.ハンプソン、ドンソン、アヌーシュカグーナワルデナ、ヴァシリスZ.マルマレリス、サムA.デッドワイラー、2011年)。 CA3およびCA1領域を包含する微小電極アレイを、脳の両側から実験動物の海馬に導入した。 ラットが手術から回復した後、一連のテストが開始されました。
ラットは、2つの格納式レバーのあるチャンバーに入れられました。 テストは3つのフェーズで構成されていました。 最初にレバーの1つがランダムに前進しました。 ラットがこのレバーを押すと、事前に選択された海馬ニューロンの活動の写真(サンプル段階)が記録されました。 その後、レバーが収縮し、あるシリーズでは1〜30秒、他のシリーズでは最大60秒の休止がありました。 その後、両方のレバーが前進しました。 元のサンプルの反対側のレバーを押すと、水滴の形で補強が行われました。 エラーが発生した場合、つまり、サンプル段階にあった同じレバーを押すと、罰が続きました-ライトを5秒間オフにします。 その結果、試行の成功の遅延時間への依存性をプロットしました(下図)。

海馬の自然活動の測定値を制御する(セオドアW.バーガー、ロバートE.ハンプソン、ドンソン、アヌーシュカグーナワルデナ、ヴァシリスZ.マルマレリス、サムA.デッドワイラー、2011年)
記録された信号に基づいて、CA3の信号に応じてCA1で予測される信号を予測する予測モデルがコンパイルされました。 一連の実験では、予測信号が既存の信号に追加されました(下図)。
人工信号の追加(Theodore W. Berger、Robert E. Hampson、Dong Song、Anushka Goonawardena、Vasilis Z.Marmarelis、Sam A.Deadwyler、2011)
興味深い結果が得られました。 結果に大きな影響を与えるのは、サンプルの提示段階で信号が調整された場合、つまり初期画像が保存された場合のみです。 残りの瞬間の信号の画像は基本的ではありませんでした。 さらに、補正信号が強いほど、最終結果は良くなります。
実験の著者は、結果を説明し、海馬が現在の情報をエンコードし、それをメモリに書き込む準備をしているという考えから始まりました。 海馬で観察される活動のパターンは、イベントの情報説明を運ぶこと。 したがって、「正しい」信号の増幅は、記憶の結果を改善するのに役立ちます。
このモデルでは、結果の説明は多少異なります。 海馬の識別子が重要なのは記憶の形成時のみであることは明らかです。 海馬によって作成されたメモリは、それを再生する必要はありません後。 海馬の現在の識別子を強化すると、暗記の力にプラスの影響がありますが、これは、より明確な情報画像を作成した結果ではなく、現在の記憶を残りの記憶に対して強調するだけです。 正常に機能している脳がこのようにすべての記憶を強調している場合、最終結果はゼロになります。
最も興味深い一連の実験は、海馬の固有の活動の抑制に関連していました。 このために、ラットに海馬のCA3領域にMK801海馬を定期的に注射しました。 MK801は、グルタミン酸を使用して正常なシナプス伝達をブロックします。 それは、グルタミン酸に敏感なNMDA受容体のイオンチャネルに侵入し、その働きを妨害します。
CA3サイトの封鎖により、結果が悪化することが予想されました。 しかし、人工信号の供給により、予想されるアクティビティの状況が復元され、正解数が大幅に改善されました(下図)。

自然な海馬信号を人工信号に置き換える(セオドア・W・バーガー、ロバート・E・ハンプソン、ドン・ソン、アヌーシュカ・グーナワルデナ、ヴァシリス・Z・マルマレリス、サム・A・デッドワイラー、2011年)
私はこの結果を、海馬で起こっていることの神経記述の奇跡的な復元と解釈し、記憶にこの記述を続けて記録するつもりはありません。 さらに、実験では、32個の電極のみを備えたマトリックスが使用され、そのうちの約半分が関与していました。 おそらく、ランダムな識別子が作成され、メモリの形成が可能になりました。
しかし、海馬ではすべてがそれほど単純ではありません。 あなたが最初に会ったときに非常に困惑しているいくつかの事実があります。 1971年、ジョン・オキーフは海馬で細胞を発見しました(オキーフ・J、ドストロフスキー・J、1971年)。 これらのセルは、内部ナビゲーターのように反応します。 ラットが長い廊下に置かれている場合、特定の細胞の活動により、ラットがどこにいるのかを正確に言うことができます。 さらに、これらの細胞の反応は、この場所に到達した方法に依存しません。
2005年に、空間位置をコードするニューロンが海馬で見つかり、座標グリッドのようなものを形成しました(Hafting T.、Fyhn M.、Molden S.、Moser MB、Moser EI、2005年)。
2011年、海馬には時間間隔を特定の方法でエンコードする細胞があることが判明しました。 彼らの活動は、他に何も起こらなくても、リズミカルなパターンを形成します(Christopher J. MacDonald、Kyle Q. Lepage、Uri T. Eden、Howard Eichenbaum、2011)。
これらすべての事実から、結論は、海馬が時間と空間の両方で私たちの位置をコーディングする責任があることを示唆しています。 同時に、海馬は兵士に似ており、「フェンスから昼食まで掘る」のは当然のことです。 海馬は、空間ナビゲーションと時系列ナビゲーションの両方を組み合わせており、偶然にも多くの共通点があります。 宇宙の旅がランドマークからランドマークへと進むように、時間の旅は時系列のマークから別のマークへと進みます。
以前、海馬は記憶の一意の識別子を形成すると主張しました。 この機能は、海馬が時空間座標を追跡するという事実とどのように関係していますか? もちろん、海馬には2つの独立したプロセスに関与する二重の機能があると仮定できます。 しかし、同じコインの両面を扱っていると仮定する方が論理的です。
一般的にどのように情報を識別するのですか? Googleを使用して、検索の意味を定義する一連の単語で構成されるクエリを作成します。 結果のリストを受け取ったら、日付または地域に制限を導入してリストを改良できます。
画像のライブラリを作成し、作成日とジオタグが付けられた場所で写真にマークを付けます。 次に、誰に何がキャプチャされているかを説明します。
ファイルをコンピューターに配置するとき、作成時間とファイルへのパスを示します。 ファイルの名前で、その内容を簡単に説明します。
最も異なるものを識別するには、一方で、それらの意味を説明する記号を使用し、他方で、任意の現象の普遍的な記号として、空間と時間で調整します。 そのような識別は、その後、ユニークなラベルとしてだけでなく、検索ツールとしても非常に便利で有用であることがわかりました。 自然は同じように進んだと考えられます。
つまり、イベントの一意の識別子を作成することにより、海馬はランダムなコードを生成して皮質全体に配布するだけでなく、あらゆる状況に最適なこのコードの時空間機能を組み込むことが理にかなっています。 このような時空間的記述自体は、さまざまな要因の組み合わせとして非常にユニークです。 それにいくつかの小さなランダムな付属物を追加することで十分であり、結果の識別子は明確に特定のイベントを示します。
このような識別は、各メモリの一意のキーよりもはるかに強力です。 さまざまな追加の関連付けの可能性。 しかし最も重要なことは、時間と空間の両方でイベントがどのように発生するかを考慮した複雑な記述をエンコードすることが可能になることです。 しかし、これについての話は少し後で行われます。
海馬の識別子が時空間座標を実際にエンコードすると仮定すると、海馬はまず、空間と時間の同様の位置を認識することを学習できるように皮質と相互作用する能力を持っていることがわかります。 第二に、その識別子を皮質の空間全体に広げるために、海馬は有限数のユニークな断片からそれらを形成しなければなりません。 これは、海馬が使用する「アルファベット」全体を配布するために皮質を事前に訓練する必要があるという事実に由来します。 新しい識別子は、「ゼロから」ではなく、既知の要素の組み合わせとして作成する必要があります。 原則として、これらの要件に複雑なことはありません。これは、自然言語のフレーズでは、すべてのフレーズが文字の有限セットから構築されることに対応しています。
中古文学
継続
前のパーツ:
パート1.ニューロン
パート2.要因
パート3.パーセプトロン、畳み込みネットワーク
パート4.バックグラウンドアクティビティ
パート5。脳波
パート6.投影システム
パート7.ヒューマンコンピューターインターフェイス
パート8.波動ネットワークの要因の分離
パート9.ニューロン検出器のパターン。 逆投影
パート10.空間的自己組織化
パート11.動的ニューラルネットワーク。 連想性
パート12.メモリーのトレース
パート13.連想メモリ
アレクセイ・レドズボフ (2014)