
この一連の記事では、従来のモデルとは大きく異なる脳の波動モデルについて説明します。 参加したばかりの人は最初の部分から読み始めることを強くお勧めします。
前のパートでは、分散メモリがどのように見えるかを示しました。 主なアイデアは、一般的な波の識別子がニューロンを結合できることであり、ニューロンはその活動によって記憶に残る画像を形成します。 特定のイベントを再現するには、皮質に沿ってメモリの対応する識別子を開始するだけで十分です。 その分布は、このメモリが修正されたときに皮質上にあった活動の同じ絵を復元します。 しかし、主な問題は、必要な識別子をどのように取得するかです。 記憶の連想性は、一連の特性に従って、これらの兆候が存在した記述内のイベントを選択できることを意味します。 つまり、特定の記号の説明に従って、これらの記号に適したメモリの識別子を取得できるようにする神経機構がなければなりません。
神経波の伝播について話したとき、ニューロンがシナプス外膜に、それが関与している波のパターンを蓄積しているという事実から始めました。 おなじみの絵に出会った後、そのスパイクをもつニューロンは独特のパターンの継続を作成します。 そしてここで重要なのは、ニューロンが波のパターンを認識するだけでなく、それ自体が伝播パターンの一部であることです。 それ自体がユニークな波の一部であることだけが、ニューロンがその伝播に参加できることです。
以降の考慮事項で混乱しないように、皮質の波動モデルの基本的な特性をもう一度繰り返しましょう。 同じ波形パターン、つまり識別子に属するニューロンに注目すると、次の図に示すポイントのセットのようなものが得られます。

皮質の任意の場所を取得し、識別子の要素をアクティブにすると、アクティブな場所から伝播する波が得られ、識別子の特徴的なパターンが繰り返されます(下図)。

皮質の各場所を通過すると、波はそのユニークなパターンの断片を「強調」します。 したがって、領域1から始まり、領域2に到達する波は、識別子によって事前に定義された独自の一意のパターンを作成します(下図)。

パターンの一意性により、皮質の各場所で、波を構成する識別子を決定できます。
領域2の穏やかな地殻で既に馴染みのあるパターンを再現すると、領域1に広がった波が作成され、そこにこの識別子の特徴である同じパターンが作成されます。

これらすべてから、識別子を認識するためには、この場所で波がどのパターンを作成するかを地殻のどこでも覚えておけば十分です。 これは、ニューロンのシナプスまたは膜のシナプス外部分で記憶できます。 シナプスでの記憶は、認識時に誘発された活動(衝動のバースト)につながり、代謝型受容クラスターでの記憶は、なじみのある波が現れたときに単一の交連を獲得することを可能にします。
識別子を再現する必要がある場合はさらに困難です。 これを行うには、必要な識別子に関連する密集したニューロンのグループの少なくとも1つのスパイクをアクティブにする必要があります。 逆投影を記述し、いくつかのメディエーターを導入して、同様の機能を実装するメカニズムを示しました。
概念の結合性について言えば、識別子の波に基づいて作成されたクエリが、クエリに含まれる概念に関連付けられた一連の概念識別子を返すことを示しました。
連想イベントメモリからメモリを抽出するメカニズムを示すために、概念記号で構成されるウェーブリクエストが、このリクエストに適したメモリの一意の海馬識別子を生成する方法を示す必要があります。 そのような識別子のセットを取得し、それらから識別子を選択して、皮質に沿ってこの識別子を実行すると、シナプス外膜にこの識別子を含むニューロン検出器のアクティビティを取得できます。これは、必要なメモリの説明図全体を復元するのと同じです。
私たちの単純化と回路図の説明について思い出させてください。 次に、脳がそのように機能すると主張せずに、連想記憶の可能な神経メカニズムの概要を説明します。
繰り返しますが、実際のニューロンの構造について簡単に説明します。 ニューロンの本体-ナマズ-は限られた領域を持ち、すべてのシナプス接触のためのスペースを提供することはできません。 ほとんどのシナプスは、樹状突起または樹状ツリーと呼ばれる分岐構造に分類されます(下図)。 樹状突起上のシナプスの数は、体細胞上のシナプスの数の10〜20倍です。
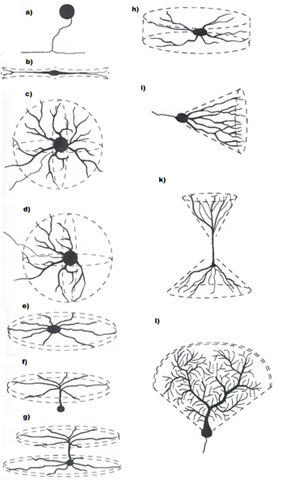
樹状の木の形態(グレッグスチュアート、ネルソンスプラストン、マイケルハウサー)
ニューロンは、信号が1つまたは異なる樹状分岐のシナプスに到達するかどうかによって異なる動作をすることがわかりました(Shepherd GM、Brayton RK、Miller JP、Segev I.、Rinzel J.、Rall W.、1985 ) 1つの分岐のシナプスにパルスが同時に到着すると、異なる分岐に分散した信号よりもはるかに強いニューロン応答が発生します。
この種の観察に基づいて、樹状分岐が一致検出器の役割を果たすことができるという仮説が生まれました(Softky、1994)。 その本質は、ニューロンのスパイクを生成するために樹状ツリーの遠隔枝で生成された信号が、体細胞への信号経路に沿って横たわっている樹状ツリーのセグメントがアクティブであることが必要であるということです。
このような効果は、ピラミッド型ニューロン(Jarsky T.、Alex Roxin A.、Kath WL、Spruston N.、2005)で示されていますが、類似の何かが他のタイプのニューロンにも特徴的であると想定できます。
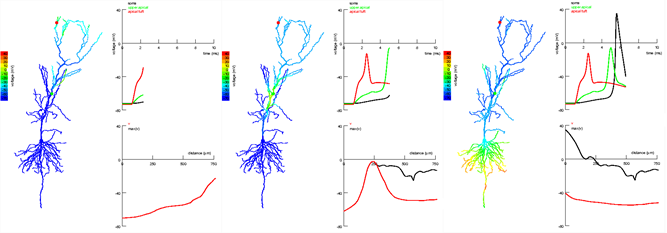
海馬錐体ニューロンの頂部および傾斜部における信号伝播位相。 信号は先端の束(赤い点)に現れ、シェーファー側副枝(緑の点)に伝搬します。 体細胞に近い樹状突起部位の信号は、遠方の信号を減衰させず、スパイクの出現に寄与します(Jarsky T.、Alex Roxin A.、Kath WL、Spruston N.、2005)
樹状突起に沿ったパルスの伝播には、その顕著な減衰が伴います。 理論的には、遠い(遠位)シナプスの効果は、近い(近位)効果よりも大幅に小さいはずです。 しかし、そのようなシナプスの寄与を均等化するメカニズムが示されており、「シナプスの民主主義」の概念をもたらしています(Clifton C. Rumsey、LF Abbott、2006)。 樹状突起分岐に沿ってシナプスの寄与を調整することにより、分岐がニューロンによって何らかの形でさらに処理される独立した論理要素と見なすことができます。 これは、理論的には、樹状突起の構造と体細胞の反応を変えて、異なる論理特性を持つニューロンを得ることができることを意味します。
たとえば、ジェフホーキンスによる「階層的時間的記憶」という一般的な概念では、「または」モードで動作する自律要素を使用するニューロンが使用されます(下図)。
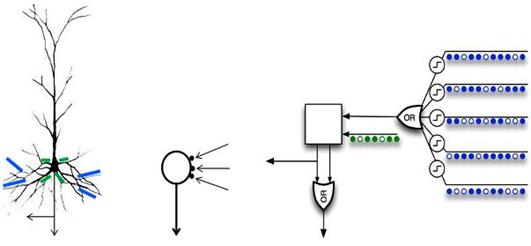
錐体ニューロンと比較した樹状突起論理自律要素のセットを備えたニューロンモデル(Hawkins、2011)
実際の脳は、非常に多様な特性を持つニューロンでも動作すると仮定することは非常に適切です。
実際、連想イベントの記憶の説明に移ります。 平らな皮質を形成する2種類のニューロンがあるとします。 これらのニューロンの受容野は、両方のタイプのニューロンが該当する環境の特定の局所領域を包含しています。 結合の相互分布を記憶しながら、条件付きで2つの層に分割します。
情報識別子の波を伝播するように、最初のタイプのニューロンを設定しましょう。 2番目のタイプのニューロンは、海馬の識別子の波のみを強制的に伝播させます(以下の図)。
異なるタイプのメディエーターを持つニューロン上の2つの独立した波の伝播
これを行うために、使用するメディエーターに応じて軸索とシナプス外受容体を希釈します(下の表)。 シナプス受容体は、波の伝播を指定するために重要ではないことに注意してください。
| シナプス | シナプスを超えて | 軸索 | |
|---|---|---|---|
| タイプ1 | A | A | |
| タイプ2 | B | B |
メディエーターとレセプターのこのようなタスクでは、2つの層の波は互いに影響しません。
次に、2番目のタイプのニューロンを複雑にし、非シナプスメディエーターによって区別される2種類のブランチからなる樹状ツリーを作成します。 シミュレーションでは、同種の枝を組み合わせることができ、その結果、「または」原理に従って機能する2つの樹状セグメントを残すことができます(下の表)。
| セグメント | シナプス | シナプスを超えて | 軸索 | |
|---|---|---|---|---|
| タイプ1 | A | A | ||
| タイプ2 | 1 | B | B | |
| 2 | A |
識別子の波の伝播方法についてニューロンをトレーニングしているときに、「外来」メディエーターに敏感な2番目のセグメントが2番目のタイプのニューロンでブロックされたとします。 2番目のタイプのニューロンでの波の形成に必要なすべての情報は、最初のセグメントのシナプス外記憶に蓄積されます。
2番目のセグメントを含めると、最初のタイプのニューロンを伝播する波のパターンを記憶し始めます。 ニューロンがスパイクを生成する瞬間にシナプス外記憶が起こると仮定します。 同時に、シナプスでの活動の瞬間的な画像は記録されませんが、波の伝播サイクル全体にわたって蓄積されます。 累積により、メモリー位置で2つのレベルの波の位相が一致しなくても、最初のレベルの波の断片を思い出すことができます。 つまり、第2レベルのニューロンがスパイクを発行すると、たとえ少し前であっても、情報アクティビティの写真をキャプチャします。
このような暗記の1つの尺度を費やしたとします。 この測定でアクティブな海馬識別波を伝播するすべての第2レベルのニューロンは、第2セグメントで、第1タイプのニューロンを通過した情報波のフラグメントを記憶します。 これは、同じ説明の波を繰り返すと、この図を認識するすべての第2レベルのニューロンがスパイクを与えることを意味します。 これらのスパイクの全体像は、この情報画像を記憶した時点での海馬のまさにその識別子によって再現されます。
そのため、さまざまな説明的な写真を記憶できます。 対応する説明を繰り返すと、第2レベルのニューロンは、現在の説明に該当する海馬のすべての識別子の完全な組み合わせを生成します。
そのような暗記は非常に無駄であることがわかります。 2番目のタイプのすべてのアクティブニューロンの各画像を記憶します。 識別子を再現するために、ウェーブをトリガーできる小さなローカルエリアのみが必要です。 状況を修正するために、2番目のタイプのすべてのアクティブニューロンではなく、1番目のタイプのニューロンの高い誘導活性を持つ領域にあるニューロンにのみ情報波を覚えます。 つまり、情報が二重であることを忘れないでください。 それは、識別波であり、誘発された活動のパターンの活動でもあります。 波は覚えておくべきパターンを与え、パターンはこの場所を示します。
場所を選択する最も簡単な方法は、パターンの空間的自己組織化に使用したのと同じ活動分野を使用することです。 この構成では、記憶は次のように表すことができます。 情報画像が誘発活動のパターンを作成する皮質の場所では、活動フィールドが形成されます。 海馬波を伝播する2番目のタイプのすべてのアクティブニューロンは、その活動フィールドが特定のしきい値を超えているため、シナプス外記憶の情報波によって記述されるパターンを修正します。
情報要求波を送信することにより、この要求を認識する2番目のタイプのニューロンのアクティビティを取得します。 このようなニューロンのローカルグループは、この要求に関連付けられているすべてのメモリに適用される海馬識別子を発行します。
メモリの一意の識別子を修正および復元する方法を示しました。 前に、ニューロン検出器がそのような一意の識別子のメモリを保存する場合、記憶された瞬間に対応する画像を復元できることを示しました。 次の表は、このようなメモリのメディエーターの可能な構成を示しています。
| セグメント | シナプス | シナプスを超えて | 軸索 | |
|---|---|---|---|---|
| タイプ1 | 1 | A | A | |
| 2 | B | |||
| タイプ2 | 1 | B | B | |
| 2 | A |
アクティビティフィールドを介して記憶領域を設定することは、かなり粗雑なツールです。 設計を改善する非常に明白な方法があります。 たとえば、2番目のタイプのニューロンのシナプストレーニングを通じて、皮質のこの場所にある概念に対応する領域をそれらから作成できます。 樹状突起一致検出器の概念を使用すると、概念の活動を、この概念に関連付けられたメモリ識別子の記録と逆生成の両方の前提条件にすることができます。
一般に、多断面ニューロンを備えた皮質の波動モデルが提供する豊富なツールにより、かなり複雑で興味深いデザインを実装できます。 物語を完全に混同しないように、すでに引用した最も単純なスキームに限定します。
中古文学
継続
前のパーツ:
パート1.ニューロン
パート2.要因
パート3.パーセプトロン、畳み込みネットワーク
パート4.バックグラウンドアクティビティ
パート5。脳波
パート6.投影システム
パート7.ヒューマンコンピューターインターフェイス
パート8.波動ネットワークの要因の分離
パート9.ニューロン検出器のパターン。 逆投影
パート10.空間的自己組織化
パート11.動的ニューラルネットワーク。 連想性
パート12.メモリーのトレース
アレクセイ・レドズボフ (2014)